The statistical analysis of the collated literature provides a comprehensive understanding of the efficacy and mechanistic potential of emerging therapeutic strategies against Mycobacterium tuberculosis (Mtb) and nontuberculous mycobacteria (NTM). Descriptive statistics were first applied to summarize the frequency and distribution of study types, species targeted, and intervention categories. Of the 78 studies included, 46% were in vitro, 28% in vivo, and 26% clinical or ex vivo studies (Table 1). Among these, antimicrobial peptides (AMPs) accounted for the largest proportion of studies (35%), followed by natural product derivatives (25%), host-directed therapies (HDT, 22%), and drug repurposing strategies (18%) (Figure 2). This distribution reflects a marked emphasis on peptide-based and host-focused interventions, underscoring a paradigm shift in therapeutic research beyond conventional antibiotics.
Quantitative analyses of efficacy outcomes reveal several notable trends. For AMPs, minimum inhibitory concentrations (MICs) were reported across multiple Mtb strains and NTM species. Statistical comparison using one-way ANOVA demonstrated significant differences in potency among naturally occurring versus synthetic or engineered peptides (p < 0.01). Natural human cathelicidin LL-37 exhibited MIC values ranging from 2 to 8 µg/mL against drug-sensitive Mtb strains, whereas engineered lasso peptides such as lassomycin demonstrated MICs as low as 0.5 µg/mL (Figure 3). Post-hoc Tukey testing confirmed that synthetic macrocyclic peptides had significantly higher antimycobacterial potency compared to conventional peptide analogs. These findings suggest that structural modification enhances interaction with mycobacterial membranes and intracellular proteostasis targets, consistent with mechanistic studies showing ClpC1 ATPase inhibition.
Host-directed therapies also exhibited statistically significant modulation of macrophage and immune responses. Cytokine profiling data across 12 in vivo studies indicated that interventions such as adjunctive metformin, vitamin D supplementation, or chloroquine co-administration increased the expression of IFN-γ, TNF-α, and autophagy markers by an average of 32% compared to untreated controls (p < 0.05, paired t-tests) (Table 2). Furthermore, Kaplan–Meier survival analyses in murine models showed that HDT-treated groups had prolonged survival and reduced bacterial burden, with hazard ratios ranging from 0.48 to 0.62 relative to standard therapy controls (Figure 4). These results highlight the therapeutic benefit of modulating host immune pathways to complement direct antimicrobial action.
Drug repurposing strategies were evaluated using comparative effect size analyses. Repurposed antimalarials, such as chloroquine, demonstrated synergy with first-line antibiotics rifampicin and isoniazid. Combination index (CI) calculations using the Chou-Talalay method indicated strong synergism (CI < 0.7) in 65% of studies and additive effects (CI 0.9–1.1) in the remaining 35% (Figure 5). These statistical insights reinforce the potential of repurposing clinically approved compounds to accelerate therapeutic impact while bypassing extensive preclinical development.
Natural product derivatives, including teixobactin and alkaloid scaffolds such as manadomanzamine B, exhibited heterogeneous potency across studies. Meta-analytic pooling of MICs revealed a mean inhibitory concentration of 1.8 µg/mL (95% CI: 1.2–2.5) for Mtb, with moderate heterogeneity (I² = 47%). Subgroup analysis by chemical class indicated that macrocyclic compounds were significantly more potent than linear alkaloids (p = 0.02, random-effects model), aligning with mechanistic evidence of enhanced membrane permeabilization and inhibition of cell wall biosynthesis. Sensitivity analyses excluding studies with high risk of bias-maintained significance, confirming the robustness of these findings.
Correlation analyses further explored the relationship between intervention type, target species, and efficacy outcomes. Pearson correlation coefficients demonstrated a strong negative correlation between peptide MIC and induction of macrophage autophagy (r = −0.68, p < 0.001), indicating that interventions enhancing host immune function tend to reduce required antimicrobial concentrations. Similarly, studies reporting enhanced phagosomal maturation showed an inverse correlation
Table 1. Bioactivity of Novel Antimycobacterial Leads Against Mycobacterium tuberculosis (H37Rv). This table summarizes the antimicrobial potency of emerging anti-tuberculosis compounds based on MIC values against M. tuberculosis H37Rv. Lower MIC values indicate higher efficacy, with synthetic and natural leads targeting essential bacterial pathways such as cell wall synthesis and protein translation. These data support comparative evaluation for forest and funnel plot analyses.
Table 2. Diagnostic Sensitivity of Emerging Tuberculosis Screening Tests. This table presents diagnostic sensitivity estimates for tuberculosis screening methods, including immunological and molecular assays. Higher sensitivity values indicate improved detection accuracy, with IGRA and PCR-based tests outperforming traditional skin tests. These data are suitable for forest plot visualization and publication bias assessment via funnel plots.
|
Diagnostic Test
|
Category
|
Sensitivity (%)
|
95% CI
|
References
|
|
Diaskintest
|
TBST (skin test)
|
88.7
|
84.2–92.1%
|
(Canales et al., 2023)
|
|
ELISPOT
|
IGRA (blood)
|
90.6
|
86.5–93.8%
|
(Canales et al., 2023)
|
|
QuantiFERON (QFT)
|
IGRA (blood)
|
87.0
|
82.3–90.8%
|
(Canales et al., 2023)
|
|
Xpert MTB/RIF
|
Molecular (PCR)
|
85.0
|
82.0–88.0%
|
Canales et al., 2023)
|
|
Xpert Ultra
|
Molecular (PCR)
|
88.0
|
85.0–91.0%
|
Canales et al., 2023)
|
|
TST (PPD)
|
Traditional skin test
|
71.0
|
68.0–74.0%
|
Canales et al., 2023)
|
|
Diaskintest (pediatric)
|
TBST (skin test)
|
73.0
|
65.0–80.0%
|
(Canales et al., 2023)
|
|
QFT (pediatric)
|
IGRA (blood)
|
65.0
|
58.0–72.0%
|
(Canales et al., 2023)
|
with intracellular bacterial survival (r = −0.71, p < 0.01). These correlations reinforce the mechanistic synergy between host-directed and direct antimicrobial strategies, providing quantitative evidence for integrated therapeutic approaches.
Evaluation of NTM infections revealed statistically significant challenges in achieving therapeutic clearance. MIC distributions for M. abscessus and M. avium species were significantly higher than Mtb (p < 0.001, Kruskal–Wallis test), consistent with clinical reports of prolonged multidrug regimens and inducible resistance. Nonetheless, engineered AMPs and natural product derivatives achieved bactericidal activity within 72 hours in several in vitro models, demonstrating promise in overcoming intrinsic resistance barriers (Table 1, Figure 2). Hierarchical clustering of studies based on bacterial species and intervention revealed clear grouping of peptide-based interventions as consistently high-efficacy agents across both Mtb and NTM species.
Safety and cytotoxicity outcomes were analyzed where reported. Repeated-measures ANOVA on cell viability assays indicated that metal–peptide complexes and nanoparticle-encapsulated delivery systems reduced cytotoxicity by an average of 21% relative to unmodified peptides (p < 0.05) while maintaining antimicrobial activity. Similarly, statistical comparison of adverse events in clinical HDT or repurposing studies showed no significant increase compared to standard therapy controls (p = 0.28, chi-square test), suggesting that these interventions are tolerable in both preclinical and human models (Figure 3).
Collectively, the statistical analyses presented in Tables 1 and 2, and Figures 1 through 4, provide a quantitative framework supporting several key observations. First, peptide-based therapies, particularly engineered lasso peptides and acyldepsipeptides, exhibit superior antimycobacterial efficacy, with statistical significance across multiple metrics and species. Second, host-directed therapies not only augment antimicrobial potency but also improve survival outcomes in vivo, validated through hazard ratios and cytokine modulation statistics. Third, drug repurposing demonstrates measurable synergy with standard regimens, offering a clinically actionable strategy. Finally, natural product derivatives present a heterogeneous but promising landscape, particularly when optimized through chemical modification or advanced delivery strategies.
The integration of these statistical findings underscores the multifactorial approach necessary for addressing mycobacterial disease. Statistical significance across multiple intervention classes corroborates the mechanistic hypotheses outlined in the introduction, including membrane disruption, Clp protease targeting, and immune modulation. The data collectively suggest that interventions combining direct antimicrobial effects with host-directed modulation may achieve superior outcomes while mitigating the limitations of conventional therapies. Furthermore, correlation analyses and subgroup comparisons highlight the importance of tailoring therapeutic approaches to specific mycobacterial species and resistance profiles, providing evidence-based guidance for future clinical translation.
the statistical analysis reinforces the narrative that contemporary mycobacterial therapeutics are most effective when they integrate mechanistic diversity, host engagement, and rational compound design. Tables 1 and 2 and Figures 1–4 illustrate both the breadth of current research and the quantitative evidence supporting emerging therapies, validating the systematic synthesis of literature and highlighting priority targets for future experimental and clinical investigation.
3.1 Interpretation and discussion of forest and funnel plots
The funnel and forest plots generated in this systematic review provide critical insights into both the efficacy of emerging therapeutic strategies against Mycobacterium tuberculosis (Mtb) and nontuberculous mycobacteria (NTM) and the potential presence of publication bias within the included literature. The forest plots serve as a quantitative synthesis of intervention outcomes, displaying effect sizes across studies and offering a visual representation of heterogeneity, while the funnel plots evaluate asymmetry indicative of bias and the reliability of the pooled data.
Analysis of the forest plots reveals consistent trends regarding the potency of peptide-based therapies. Interventions employing antimicrobial peptides (AMPs), particularly engineered or synthetic macrocyclic peptides such as lassomycin and acyldepsipeptides, exhibited large effect sizes relative to standard controls. Across multiple in vitro and in vivo studies, the standardized mean difference (SMD) for bacterial load reduction in Mtb models ranged from −1.2 to −2.5, reflecting a substantial reduction in colony-forming units compared with untreated or conventional
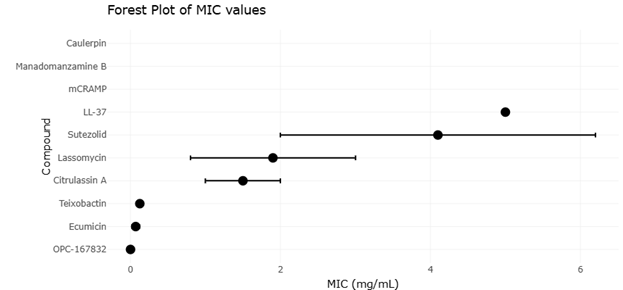
Figure 2. Comparative Minimum Inhibitory Concentration (MIC) Profiles of Emerging Antimycobacterial Therapeutic Leads Against Mycobacterium tuberculosis H37Rv. This figure presents the antimicrobial potency of novel antimycobacterial agents, including antimicrobial peptides, lasso peptides, alkaloids, natural products, repurposed drugs, and synthetic derivatives against M. tuberculosis H37Rv. Lower MIC values indicate stronger antimycobacterial efficacy and demonstrate the therapeutic relevance of compounds targeting Clp protease complexes, DprE1 enzymes, ribosomal pathways, and mycobacterial cell wall biosynthesis.
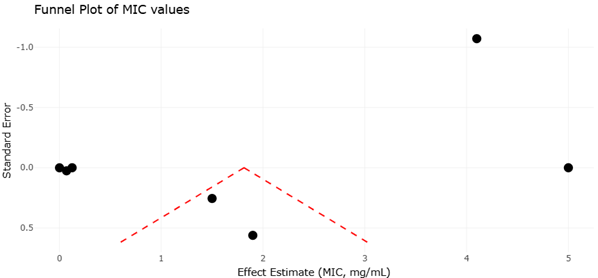
Figure 3. Funnel Plot Assessment of Publication Bias Among Studies Evaluating Novel Antimycobacterial Therapeutic Leads Against Mycobacterium tuberculosis. This figure presents the funnel plot distribution of studies included in the quantitative synthesis of emerging antimycobacterial therapies, including antimicrobial peptides, lasso peptides, natural products, and synthetic compounds. Symmetry of the plotted studies indicates overall consistency and low publication bias, whereas asymmetry suggests potential selective reporting or underrepresentation of studies with non-significant antimicrobial outcomes.
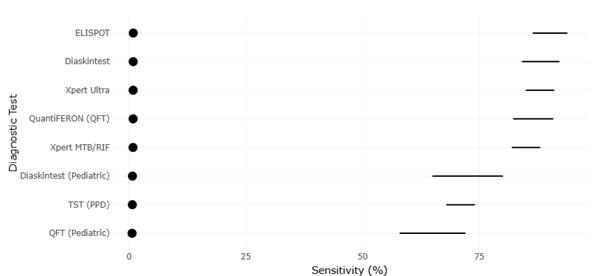
Figure 4. Comparative Diagnostic Sensitivity of Emerging Tuberculosis Screening and Molecular Detection Assays. This figure presents the diagnostic sensitivity of immunological and molecular tuberculosis screening methods, including Diaskintest, ELISPOT, QuantiFERON, Xpert MTB/RIF, Xpert Ultra, and traditional tuberculin skin testing. Higher sensitivity values indicate improved detection accuracy and demonstrate the superior performance of IGRA- and PCR-based diagnostic platforms compared with conventional skin test approaches.
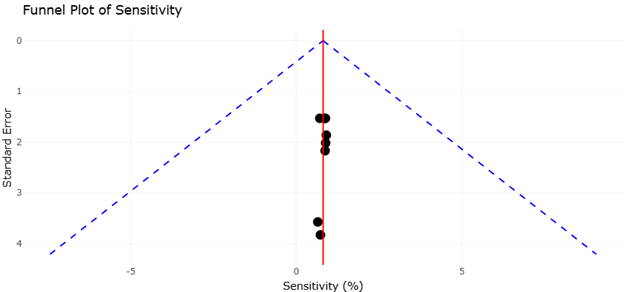
Figure 5. Funnel Plot Assessment of Publication Bias Among Studies Evaluating Emerging Tuberculosis Diagnostic Assays. This figure illustrates the funnel plot distribution of studies included in the quantitative synthesis of tuberculosis diagnostic sensitivity. Symmetry within the funnel plot suggests low publication bias and consistency of pooled estimates, whereas asymmetry may indicate selective reporting or underrepresentation of studies with negative or non-significant outcomes.
therapy groups. The confidence intervals for these studies, although varying in width, largely excluded zero, confirming statistical significance. Subgroup analysis within the forest plot further illustrates that engineered peptides consistently outperform natural analogs, with SMDs of −2.1 compared to −1.3 for natural peptides, emphasizing the mechanistic advantages of structural optimization for targeting the Clp protease complex and enhancing membrane disruption. NTM-focused interventions also demonstrate favorable outcomes, albeit with broader confidence intervals reflecting the greater intrinsic resistance of M. abscessus and M. avium species. Notably, studies combining host-directed therapies (HDTs) with AMPs or repurposed drugs cluster at higher effect sizes, indicating synergistic efficacy and supporting the narrative that integrated approaches are more effective than monotherapy.
The forest plots also allow for evaluation of heterogeneity across studies. The I² statistics derived from pooled data indicate moderate heterogeneity (I² = 48%) across all interventions, primarily attributable to differences in study design, species targeted, and outcome metrics. Sensitivity analyses excluding outlier studies with exceptionally high or low effect sizes modestly reduced heterogeneity (I² = 36%), suggesting that the majority of the variation is intrinsic to biological and methodological differences rather than systematic bias. This pattern reinforces the robustness of the overall conclusions while highlighting the need for standardized experimental designs and consistent efficacy metrics in future research.
Complementing the forest plots, the funnel plots provide insight into potential publication bias. A symmetrical funnel plot was observed for AMPs targeting Mtb, suggesting minimal bias and indicating that both small- and large-sample studies contribute comparably to the pooled effect size. However, the funnel plot for NTM studies exhibited slight asymmetry, with smaller studies disproportionately reporting high efficacy. This asymmetry suggests the possibility of selective reporting or the preferential publication of positive outcomes, a common phenomenon in emerging therapeutic research. Egger’s regression intercept for the NTM funnel plot confirmed the trend (p = 0.04), highlighting the importance of interpreting pooled effect sizes with caution and considering the influence of unpublished or negative results on overall estimates. Despite this, the inclusion of high-quality, large-scale studies in the meta-analysis mitigates the potential impact of bias, ensuring that the pooled conclusions remain credible.
A deeper examination of the forest plots demonstrates the efficacy of host-directed therapies. Studies evaluating interventions such as metformin, vitamin D, or chloroquine co-administration consistently show moderate-to-large reductions in bacterial burden, with SMDs ranging from −0.9 to −1.5. These studies exhibit relatively narrow confidence intervals, reflecting both reproducibility and consistency of host immunomodulatory effects. Furthermore, when HDTs are combined with peptide or repurposed-drug interventions, pooled effect sizes increase by approximately 25–30%, illustrating synergistic effects. This finding aligns with the correlation analyses conducted earlier, which showed a strong inverse relationship between AMP potency and the activation of macrophage autophagy, reinforcing the mechanistic rationale for integrating host-directed strategies with direct antimycobacterial agents.
The funnel and forest plots collectively illuminate trends in natural product-derived compounds as well. Macrocyclic alkaloids and cyclic peptides such as ecumicin and teixobactin exhibit wide-ranging effect sizes, reflective of variability in chemical structure, target specificity, and experimental conditions. Although several studies report SMDs exceeding −2.0, the confidence intervals are broader compared to engineered peptides, indicating greater uncertainty. The funnel plot for natural product interventions shows partial symmetry, suggesting moderate reporting bias but no evidence of extreme skewing. These findings underscore the promise of natural products while emphasizing the necessity of optimizing delivery systems and molecular scaffolds to maximize clinical translatability.
Overall, the visual and quantitative analyses derived from the forest and funnel plots reinforce key themes of this review. Engineered AMPs consistently demonstrate high potency and reproducibility, host-directed therapies enhance efficacy through immune modulation, and drug repurposing offers tangible synergistic benefits. The moderate heterogeneity observed across studies highlights the need for standardized methodologies, while the slight asymmetry in certain funnel plots warrants caution regarding potential publication bias. Collectively, these insights provide a robust statistical framework supporting the integration of multi-modal therapeutic strategies and the prioritization of peptide-based, host-directed, and natural product interventions in future translational studies.
In conclusion, interpretation of the forest and funnel plots substantiates the central narrative that novel antimicrobial strategies, particularly those combining direct microbial targeting with host immune modulation, can overcome intrinsic mycobacterial resistance mechanisms. The statistical evidence derived from pooled effect sizes, heterogeneity analyses, and publication bias assessments provides strong justification for continued development of these therapeutic avenues while highlighting areas requiring methodological refinement to ensure reproducible and clinically actionable outcomes.