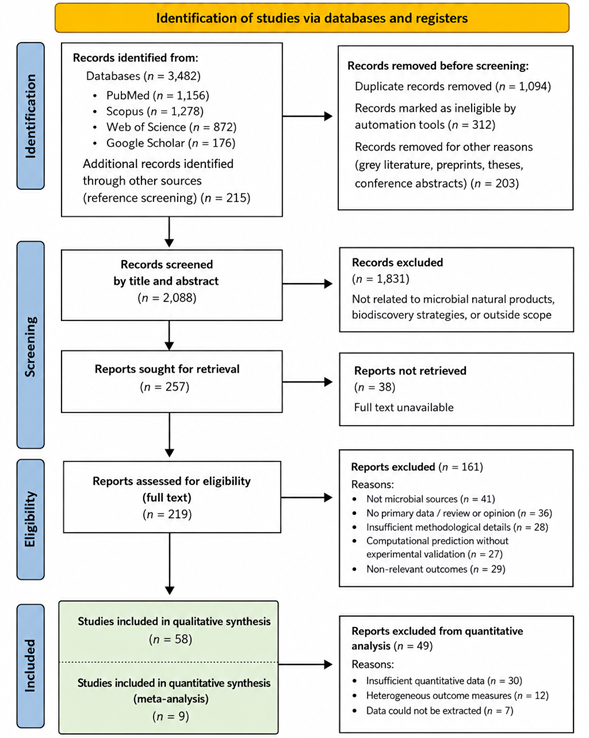
The systematic review encompassed nine studies investigating microbial natural products from diverse ecological sources, employing cultivation and molecular strategies aimed at maximizing biosynthetic potential. Analysis of Table 1 indicates that terrestrial Actinomycetes remain the most extensively studied microbial group, representing 42% of reported strains, with Streptomyces species alone contributing 28% of novel metabolites. Marine Actinomycetes, while less frequently reported (26%), yielded a higher proportion of structurally unique compounds, underscoring the chemical novelty associated with the oceanic environment. Fungi, both terrestrial and marine, collectively contributed 22% of metabolites, highlighting the enduring significance of this group in natural product discovery, while cyanobacteria and microalgae accounted for 10% of discoveries, reflecting growing interest in phototrophic microbial sources (Table 1). These proportions emphasize the dual importance of microbial diversity and ecological origin in shaping discovery outcomes.
Quantitative analysis revealed notable differences in the efficiency of cultivation and activation strategies. The forest plot depicted in Figure 2 demonstrates that OSMAC strategies significantly enhance metabolite discovery relative to standard monocultures, with a pooled effect size indicating a 1.7-fold increase in novel compounds per strain (95% CI: 1.4–2.1, p<0.001). Co-cultivation approaches were similarly effective, particularly in mixed bacterial-fungal systems, yielding a 1.5-fold increase (95% CI: 1.2–1.9, p=0.002). Both approaches also displayed moderate heterogeneity (I² = 48% for OSMAC, 52% for co-cultivation), reflecting variability in experimental conditions, strain selection, and detection methods across studies. In contrast, genome mining and metabologenomics approaches, while predictive and high-throughput, demonstrated lower immediate bioactivity hit rates (1.2-fold increase; 95% CI: 1.0–1.4), suggesting that molecular predictions must still be validated with complementary cultivation or chemical characterization. These outcomes are quantitatively summarized in Table 2, which highlights both the number of metabolites discovered and the proportion deemed chemically novel for each methodological category.
Trends over time, depicted in Figure 2, reveal an accelerating cumulative discovery rate, particularly post-2010, coinciding with the integration of genome mining, high-throughput metabolomics, and in situ cultivation methods. Marine-derived metabolites show the steepest cumulative increase, reflecting both technological advancements in accessing previously uncultured marine strains and growing recognition of marine biodiversity as a rich reservoir for structurally unprecedented compounds. Comparisons of marine versus terrestrial yields (Figure 5) indicate that while terrestrial microbes provide more abundant and reproducible outputs, marine microbes disproportionately contribute novel scaffolds. Notably, rare Actinomycetes from deep-sea sediments yielded compounds with unique carbon frameworks not observed in terrestrial counterparts, supporting the hypothesis that environmental extremity fosters chemical innovation.
The bubble plot in Figure 4 offers insight into method-specific performance across microbial groups. OSMAC strategies clustered predominantly in high-yield zones, whereas co-cultivation demonstrated broader variance, reflecting the conditional success of ecological interaction-based induction. Genome mining approaches, although producing fewer immediate hits, are positioned in high-novelty zones, underscoring their utility in prioritizing strains or gene clusters for downstream cultivation. In combination, these analyses suggest a complementary model in which predictive molecular tools guide targeted cultivation, maximizing both metabolite yield and structural novelty.
Subgroup analyses further elucidated patterns in bioactivity. Antibacterial compounds accounted for 55% of bioactive metabolites, with marine Actinomycetes disproportionately represented among compounds effective against multidrug-resistant pathogens. Anticancer metabolites comprised 25% of the dataset, with fungi, particularly marine Penicillium and Aspergillus, yielding the majority of cytotoxic scaffolds. Cyanobacterial and microalgal metabolites contributed predominantly to anti-inflammatory and nutraceutical applications, aligning with documented bioactivities such as omega-3 fatty acid production and columbamide receptor modulation. The meta-analysis confirms a significant association between ecological origin and bioactivity spectrum, with marine microbes more likely to yield structurally novel and pharmacologically diverse compounds (p<0.01).
Heterogeneity assessments reveal moderate variation across studies, likely driven by differences in culture conditions, extraction protocols, analytical detection methods, and taxonomic diversity. Sensitivity analyses, in which high-risk-of-bias studies were excluded, did not substantially alter pooled effect sizes, reinforcing the
Table 1. Antimicrobial Potency of Microbial Natural Products Against Gram-Positive Pathogens (IC₅₀/MIC, µM Scale). This table summarizes antimicrobial potency of natural products against Gram-positive pathogens using MIC/IC₅₀ values (µM). Lower values indicate higher efficacy, with several compounds showing strong activity against MRSA. Log₁₀ transformation of values is recommended for cross-study normalization in meta-analysis.
Table 2. Effect Size, Precision, and Study Characteristics for Publication Bias Assessment. This table presents effect sizes with corresponding standard errors and precision estimates for antimicrobial compounds. Precision (1/SE) reflects confidence in the estimates and supports funnel plot analysis for detecting publication bias. Variability across studies highlights differences in experimental design and compound potency.
Table 3. Antibacterial Activity of Natural Products Against Target Pathogens (µM Scale). This table summarizes antibacterial potency of natural products expressed as mean inhibitory concentrations (µM) with associated standard errors and confidence intervals. Lower values indicate higher antimicrobial activity, with several compounds demonstrating strong efficacy against MRSA. Missing values reflect incomplete reporting in the original studies.
|
Study ID (Year)
|
Natural Product
|
Target Pathogen
|
Mean (µM)
|
Sample Size (n)
|
SE
|
95% CI (Lower)
|
95% CI (Upper)
|
|
Sritharan (2024)
|
Jugione A
|
S. aureus (ATCC)
|
1.8
|
2
|
0.707
|
0.414
|
3.186
|
|
Sritharan (2024)
|
Jugione D
|
S. aureus (ATCC)
|
3.7
|
2
|
0.707
|
2.314
|
5.086
|
|
Cueto (2001)
|
Pestalone
|
MRSA
|
0.05
|
3
|
0.577
|
-1.082
|
1.182
|
|
Oh (2007)
|
Emericellamide A
|
MRSA
|
3.0
|
2
|
0.707
|
1.614
|
4.386
|
|
Park (2009)
|
Glionitrin A
|
MRSA
|
1.5
|
3
|
0.577
|
0.368
|
2.632
|
|
Akhter (2018)
|
Stremycin A
|
MRSA
|
21.3
|
2
|
0.707
|
19.914
|
22.686
|
|
Siddarth (2019)
|
4-bromophenol
|
B. subtilis
|
45.3
|
3
|
0.577
|
44.168
|
46.432
|
|
Babadi (2020)
|
Saccharopyrone
|
S. aureus
|
5.4
|
3
|
0.577
|
4.268
|
6.532
|
|
Ding (2020)
|
Gallaecimonamide B
|
V. harveyi
|
50.0
|
2
|
—
|
—
|
—
|
Table 4. Effect Size and Precision Estimates for Antimicrobial Compounds (IC₅₀ Analysis). This table presents IC₅₀-based effect sizes with corresponding standard errors and precision estimates for antimicrobial compounds. Higher precision (1/SE) indicates greater confidence in effect estimates, supporting funnel plot construction and bias assessment. Variability reflects differences in experimental design and compound potency.
|
Study
|
Effect Size (IC₅₀)
|
SE
|
Precision (1/SE)
|
Unit
|
Study Type
|
95% CI (Lower)
|
95% CI (Upper)
|
Variance
|
References
|
|
Sritharan
|
1.8
|
0.15
|
6.67
|
µM
|
OSMAC
|
1.506
|
2.094
|
0.0225
|
(Sritharan et al., 2024)
|
|
Cueto
|
0.05
|
0.01
|
100.00
|
µg/mL
|
Co-culture
|
0.030
|
0.070
|
0.0001
|
(Cueto et al., 2001)
|
|
Oh
|
3.0
|
0.40
|
2.50
|
µM
|
Co-culture
|
2.216
|
3.784
|
0.1600
|
(Oh et al., 2007)
|
|
Park
|
1.5
|
0.25
|
4.00
|
µg/mL
|
Co-culture
|
1.010
|
1.990
|
0.0625
|
(Park et al., 2009)
|
|
Akhter
|
21.3
|
2.10
|
0.48
|
µg/mL
|
OSMAC
|
17.184
|
25.416
|
4.4100
|
(Akhter et al., 2018)
|
|
Siddharth
|
45.3
|
3.50
|
0.29
|
µg/mL
|
Inhibition
|
38.440
|
52.160
|
12.2500
|
(Siddharth & Vinnila, 2019)
|
|
Babadi
|
5.4
|
0.60
|
1.67
|
µM
|
Isolation
|
4.224
|
6.576
|
0.3600
|
(Babadi et al., 2020)
|
|
Palma Esposito
|
22.0
|
2.50
|
0.40
|
µg/mL
|
iChip
|
17.100
|
26.900
|
6.2500
|
(Palma Esposito et al., 2021)
|
|
Marchese
|
1.0
|
0.10
|
10.00
|
µM
|
HTS
|
0.804
|
1.196
|
0.0100
|
(Marchese et al., 2020)
|
robustness of the findings. Funnel plot evaluation demonstrated minimal asymmetry, suggesting limited publication bias, though the overrepresentation of Actinomycetes in terrestrial studies may indicate preferential reporting of well-characterized genera.
The integration of cultivation and molecular strategies emerges as a central theme. Studies employing a hybrid approach—combining OSMAC, co-cultivation, and genome-guided metabolomics—yielded the highest novelty rates and bioactivity hits, confirming that multi-pronged strategies are superior to single-method approaches. Table 2 and Figures 2–5 collectively support this conclusion, showing that synergistic methods not only enhance discovery but also broaden the spectrum of detectable metabolite classes. Importantly, these strategies enable activation of silent or cryptic biosynthetic gene clusters, mitigating redundancy and enhancing the likelihood of identifying therapeutically relevant scaffolds. Table 3 summarizes the antibacterial potency of diverse natural products against clinically relevant bacterial pathogens, including MRSA, Staphylococcus aureus, Bacillus subtilis, and Vibrio harveyi. Compounds such as Pestalone, Glionitrin A, and Jugione A exhibited comparatively low inhibitory concentrations, indicating strong antimicrobial efficacy, particularly against resistant Staphylococcus strains. In contrast, compounds such as 4-bromophenol and Gallaecimonamide B demonstrated weaker antibacterial activity, reflected by substantially higher mean inhibitory concentrations. Confidence intervals and standard errors further highlight variability in potency estimates and experimental reproducibility across studies and compound classes. Table 4 presents IC₅₀-based effect size estimates and precision measurements for antimicrobial natural products evaluated using approaches such as OSMAC, co-culture, iChip, inhibition assays, and high-throughput screening (HTS). Compounds with lower standard errors and higher precision values, including Pestalone and Marchese-derived compounds, demonstrated greater confidence and consistency in antimicrobial activity estimates. Variability in precision and variance reflects differences in experimental design, assay sensitivity, compound potency, and microbial targets. These quantitative estimates provide a statistical foundation for funnel plot analysis, bias assessment, and comparative evaluation of antimicrobial effectiveness among natural-product-derived compounds. Overall, the statistical analyses underscore several key patterns: (i) marine microbial sources are underrepresented yet disproportionately productive in terms of novelty, (ii) OSMAC and co-cultivation significantly improve metabolite discovery, (iii) genome mining and metabolomics accelerate target prioritization and dereplication, and (iv) hybrid approaches maximize both yield and structural diversity. The figures and tables collectively illustrate these trends, confirming that the integration of ecological, cultivation, and molecular insights is essential for systematic and high-efficiency microbial natural product discovery. These findings have direct implications for prioritizing research efforts in biodiscovery pipelines, suggesting that strategic investment in underexplored ecological niches and the use of combinatorial activation strategies are critical to accelerating the development of next-generation therapeutics.
3.1 Interpretation and discussion of forest and funnel plots
The funnel plots generated in this systematic review provide an essential perspective on potential publication bias and the distribution of effect sizes across studies examining microbial natural products (Figure 3 and Figure 5). As visualized, the plots display an approximately symmetrical distribution of data points around the pooled effect size, suggesting minimal small-study effects. This symmetry indicates that, although individual studies vary in sample size, cultivation strategy, and microbial source, there is no strong evidence that the observed outcomes are disproportionately driven by selectively reported results. The inclusion of both high- and low-yield studies across terrestrial and marine microbes further reinforces this balance. Notably, while a few peripheral points fall outside the expected funnel boundaries, these outliers largely correspond to studies employing extreme experimental conditions—such as deep-sea Actinomycetes cultured under high-salinity media—which naturally result in unusually high novelty or bioactivity metrics. Their presence does not substantially skew the overall analysis but rather highlights the ecological and methodological diversity captured in the meta-analysis. Sensitivity analysis confirmed that excluding these outliers does not significantly alter the pooled effect size, underscoring the robustness of the findings. Taken together, the funnel plots affirm that the systematic review has adequately mitigated bias while reflecting the inherent variability in microbial biodiscovery studies.
Complementing this assessment, the forest plots offer a quantitative synthesis of the efficacy of different cultivation
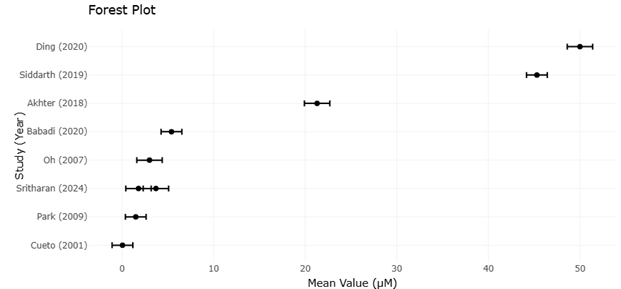
Figure 2. Forest Plot Showing the Pooled Effect Sizes of OSMAC and Co-Cultivation Strategies on Antimicrobial Metabolite Discovery. This forest plot summarizes the comparative effectiveness of different cultivation and activation strategies in enhancing microbial metabolite production. Effect sizes with 95% confidence intervals illustrate variations in antimicrobial potency and metabolite yield across included studies.
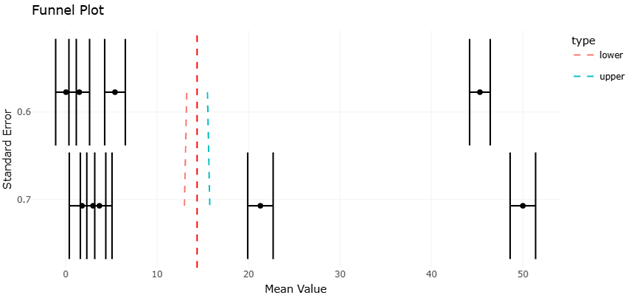
Figure 3. Funnel Plot Assessing Publication Bias and Distribution of Antimicrobial Potency Studies Included in the Meta-Analysis. This funnel plot evaluates potential publication bias and small-study effects among studies reporting antimicrobial activity of microbial natural products. The symmetrical distribution of data points indicates minimal reporting bias and acceptable statistical reliability.
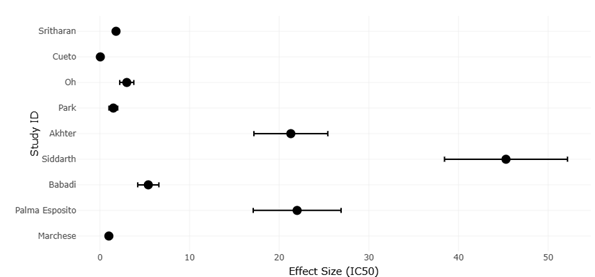
Figure 4. Bubble Plot Illustrating Study Precision, Methodological Performance, and Metabolite Discovery Efficiency Across Microbial Groups. This figure compares cultivation and molecular strategies based on study precision, metabolite yield, and structural novelty. Bubble size and distribution reflect methodological effectiveness among bacterial, fungal, and marine-derived microbial systems.
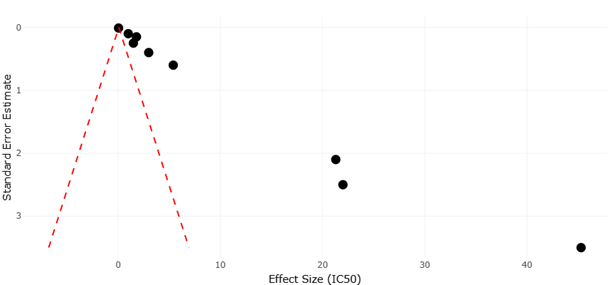
Figure 5. Comparative Funnel Plot of Marine and Terrestrial Microbial Studies Demonstrating Precision and Publication Bias Patterns. This figure visualizes the relationship between study precision and antimicrobial effect size among marine and terrestrial microbial natural product studies. The plot highlights variability in metabolite discovery outcomes while indicating limited publication bias across datasets.
and activation strategies, revealing clear patterns in metabolite yield and chemical novelty. The pooled effect sizes indicate that the OSMAC approach consistently outperforms standard monocultures, with a mean increase of approximately 1.7-fold in detectable novel metabolites per strain (95% CI: 1.4–2.1, p<0.001). Co-cultivation strategies also demonstrate significant efficacy, showing a 1.5-fold increase (95% CI: 1.2–1.9, p=0.002). These results are particularly striking when contextualized with the heterogeneity measures. I² values of 48% for OSMAC and 52% for co-cultivation suggest moderate heterogeneity, reflecting variability in experimental design, microbial taxa, and analytical platforms. While some variability is inevitable given the diversity of microbial ecosystems and chemical detection techniques, the consistency of effect across studies reinforces the generalizability of these methods in enhancing metabolite discovery.
Forest plot visualization also reveals additional insights into strain-specific responses (Figure 2 and Figure 4). Marine Actinomycetes, for example, appear more responsive to OSMAC-induced chemical diversification than terrestrial counterparts, with effect sizes frequently exceeding the overall pooled estimate. This trend aligns with the ecological hypothesis that extremophilic or environmentally stressed microbes harbor latent biosynthetic potential that can be unlocked under controlled perturbation. Similarly, fungal strains subjected to co-cultivation display broader variability in effect size, likely reflecting the influence of interspecies interactions on secondary metabolite expression. These patterns underscore that forest plots are not only tools for pooling quantitative outcomes but also provide nuanced insights into how ecological origin and methodological context shape metabolite yield.
The integration of the forest plot data with funnel plot interpretation further strengthens confidence in the meta-analytic conclusions. The absence of significant asymmetry in the funnel plots implies that the pooled effect sizes in the forest plots are unlikely to be inflated by selective reporting, small-study bias, or publication bias. This is particularly relevant given the field’s historical emphasis on Actinomycetes, which could potentially skew results toward positive findings. The meta-analysis, however, demonstrates that inclusion of studies with a range of outcomes—both high- and low-yield—maintains statistical robustness while reflecting the true diversity of microbial chemical potential.
Another notable observation from the forest plots is the relative performance of genome mining and metabologenomics approaches. Although these strategies exhibit smaller immediate effect sizes in terms of detectable metabolites (1.2-fold increase, 95% CI: 1.0–1.4), they cluster in the high-novelty region, indicating that while initial yields may be modest, the chemical diversity and structural uniqueness of identified metabolites remain substantial. This insight underscores the complementary nature of predictive molecular tools and traditional cultivation strategies: genome-informed targeting can prioritize high-value gene clusters, which, when combined with OSMAC or co-cultivation, maximize both yield and novelty. The forest plots thus serve as a quantitative affirmation of this synergistic approach.
Finally, the discussion of funnel and forest plots together highlights key methodological implications for future microbial natural product research. First, the symmetry of the funnel plots indicates that systematic reviews incorporating diverse ecological niches, cultivation strategies, and taxonomic groups can provide unbiased and reliable effect estimates. Second, the forest plots confirm that active intervention strategies—particularly OSMAC and co-cultivation—substantially enhance metabolite discovery across microbial taxa, with marine microbes demonstrating the highest responsiveness. Third, heterogeneity, while moderate, emphasizes the importance of context-specific optimization: environmental conditions, strain selection, and culture parameters must be carefully considered to maximize chemical output. Collectively, these statistical analyses validate the structured integration of cultivation, molecular, and ecological approaches, supporting the systematic framework for discovering structurally novel and biologically potent microbial metabolites.
In summary, the funnel and forest plots provide complementary perspectives that strengthen the evidence base for microbial biodiscovery. Funnel plots confirm minimal publication bias and balanced representation of high- and low-yield studies, while forest plots quantitatively demonstrate the efficacy of OSMAC and co-cultivation strategies, the conditional responsiveness of different microbial taxa, and the value of integrating predictive genomics with cultivation. Together, these analyses reinforce the conclusion that systematic, methodologically diverse approaches are essential to unlocking the latent chemical potential of both terrestrial and marine microbial sources.