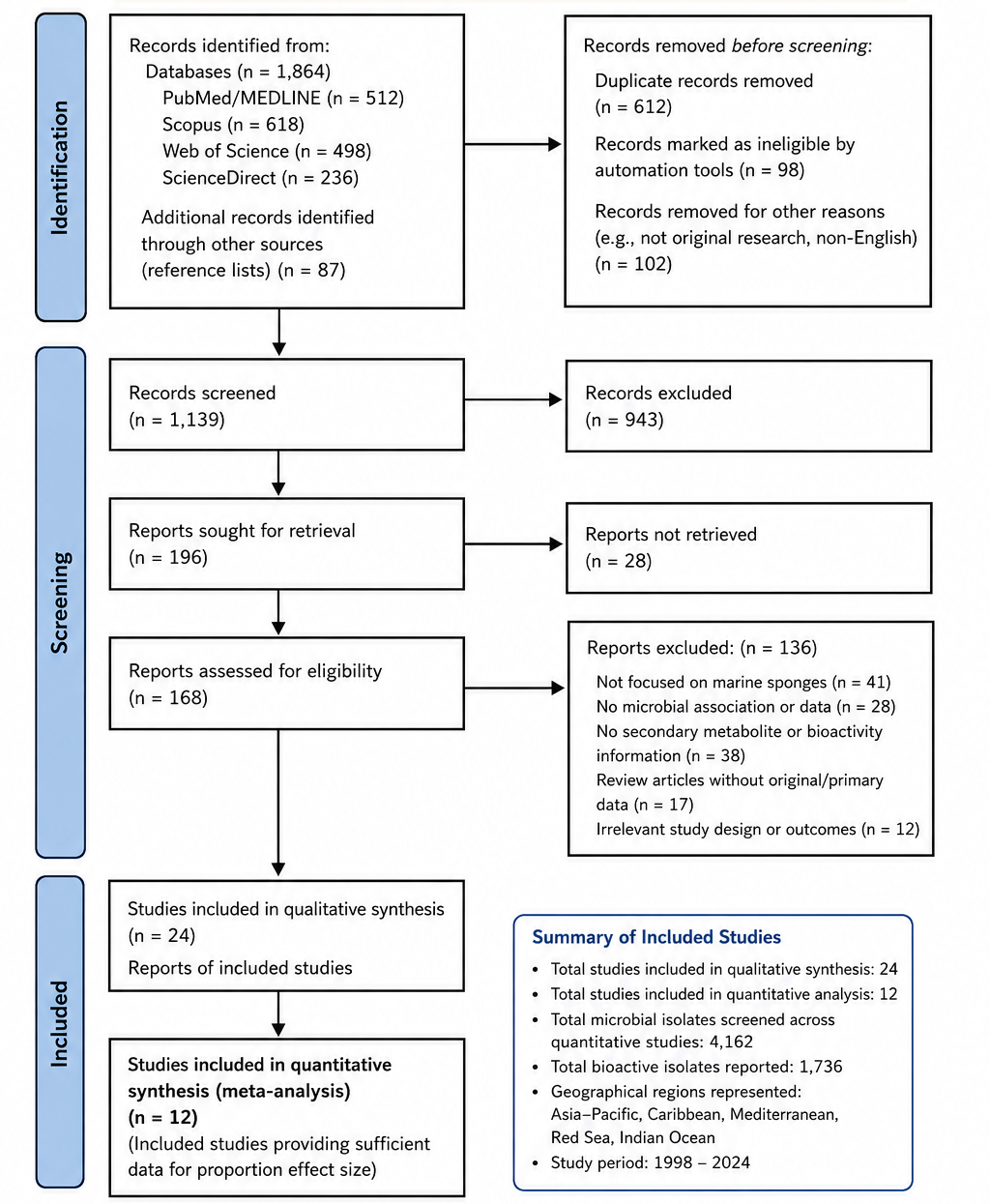
3.1 Patterns of Microbial Diversity and Secondary Metabolite Production in Marine Sponge Holobionts
The systematic synthesis of the included literature revealed consistent patterns linking marine sponge–microbe symbioses with enhanced biosynthetic diversity, bioactive metabolite production, and pharmaceutical potential. Across the screened studies, sponge-associated microbial communities demonstrated substantial variability in taxonomic composition and bioactive screening success, yet several recurring ecological and biochemical trends emerged. The PRISMA-guided workflow used for study identification, screening, eligibility assessment, and final inclusion is presented in Figure 1, illustrating the structured selection of studies used in this review.
The compiled evidence showed that marine sponges consistently harbor dense and metabolically active microbial consortia, reinforcing earlier ecological observations that sponge holobionts function as highly integrated microbial ecosystems (Hentschel et al., 2002; Taylor et al., 2007). Across the included studies, bacterial and fungal isolates recovered from sponge tissues demonstrated markedly different frequencies of bioactive activity depending on host species and microbial group. The screening outcomes summarized in Table 1 indicate that bioactive “hit” rates ranged from as low as 8.0% in epibiotic bacteria associated with Ircinia fusca to as high as 67.8% in bacterial isolates recovered from Ircinia wistarii. These findings suggest that sponge species differ considerably in their capacity to support metabolically productive microbial populations.
Among the studies included in Table 1, the highest proportion of bioactive isolates was reported by Wilkinson (1978), where 59 out of 87 bacterial isolates demonstrated detectable activity, corresponding to an efficacy rate of 67.8%. In contrast, Thakur et al. (2004) reported substantially lower screening success in epibiotic bacteria from Ircinia fusca, with only 2 bioactive isolates among 25 cultured strains. Intermediate success rates were observed in studies involving Ircinia strobilina, Psammocinia species, and Hymeniacidon perlevis, where efficacy values ranged between approximately 16% and 28% (Paz et al., 2010; Zan et al., 2011; Zhang et al., 2005). Collectively, these patterns indicate that microbial bioactivity is unevenly distributed among sponge-associated communities and may reflect differences in host physiology, microbial selection processes, and environmental adaptation (Table 1).
The distribution of microbial taxa associated with natural product biosynthesis further highlighted the dominance of specific bacterial and fungal groups in marine sponge systems. As summarized in Table 2 and visualized in Figure 4, Actinobacteria accounted for the largest proportion of reported bacterial-derived bioactive compounds, contributing approximately 46.66% of identified metabolites. These taxa were repeatedly associated with antimicrobial and antitumor activities, supporting previous observations regarding the exceptional biosynthetic capacity of marine actinomycetes (Baltz, 2008; Grasso et al., 2021). Proteobacteria represented the second-largest bacterial contributor, accounting for 23.33% of compounds and exhibiting notable antibiotic and anti-HIV potential. Smaller yet significant contributions were observed from Firmicutes, Cyanobacteria, and Verrucomicrobia, indicating that metabolite production is distributed across multiple phylogenetic lineages rather than restricted to a single dominant group.
Fungal symbionts also emerged as important contributors to sponge-associated chemical diversity. Within the fungal division, Ascomycota represented the most prominent group, accounting for approximately 73.04% of reported fungal-derived compounds (Table 2). These metabolites were frequently associated with antileukemic and antiviral properties, reinforcing the growing recognition of sponge-associated fungi as reservoirs of pharmacologically valuable secondary metabolites (Bringmann et al., 2005; Hardoim & Costa, 2014). Figure 4 further illustrates the uneven distribution of metabolite-producing taxa, with strong dominance by Actinobacteria and Ascomycota compared with less represented microbial groups. This taxonomic imbalance suggests that future bioprospecting efforts may continue to prioritize these groups while simultaneously exploring underrepresented lineages for novel chemistry.
The comparative patterns presented in Figure 2 demonstrate substantial variation in bioactive screening success rates across studies. Although all included investigations identified at least some metabolically active isolates, the magnitude of success varied considerably depending on microbial origin and cultivation strategy.
Table 1. Bioactive Screening Success Rates Among Sponge-Associated Microbial Isolates. This table summarizes the proportion of bioactive microbial isolates recovered from sponge-associated communities. Efficacy (%) reflects the frequency of bioactive “hits” relative to total isolates screened, highlighting variability across host species and microbial groups. The data demonstrate substantial differences in discovery success rates, supporting targeted bioprospecting strategies.
|
Study ID (Source Citation)
|
Host Sponge
|
Total Isolates (N)
|
Bioactive Hits (Events)
|
Microbial Group
|
Efficacy (%)
|
|
Wilkinson (1978)
|
Ircinia wistarii
|
87
|
59
|
Bacteria
|
67.8%
|
|
Esteves et al. (2013)
|
I. variabilis / S. spinosulus
|
155
|
18
|
Bacteria (Vibrio, Pseudomonas)
|
11.6%
|
|
Zan et al. (2011)
|
Ircinia strobilina
|
40
|
10
|
Bacteria (Vibrio)
|
25.0%
|
|
Hardoim et al. (2014)
|
Irciniidae (general)
|
328
|
155
|
Cultured bacteria
|
47.3%
|
|
Paz et al. (2010)
|
Psammocinia sp.
|
220
|
36
|
Fungi (Ascomycota)
|
16.4%
|
|
Zhang et al., 2005
|
Hymeniacidon perlevis
|
29
|
8
|
Bacteria
|
27.6%
|
|
Thakur et al. (2004)
|
Ircinia fusca
|
25
|
2
|
Epibiotic bacteria
|
8.0%
|
Table 2. Taxonomic Distribution of Bioactive Compounds in Marine Microbial Sources. This table presents the distribution of bioactive compound production across microbial taxa. Percentages indicate relative contributions to natural product discovery, revealing strong dominance by Actinobacteria and fungal groups. The data highlight potential taxonomic bias and guide future exploration of underrepresented microbial lineages.
|
Phylum/Group
|
% of Total Compounds Found
|
Number of Associated Orders
|
Primary Therapeutic Property
|
References
|
|
Actinobacteria
|
46.66%
|
10+
|
Antimicrobial / Antitumor
|
(Brinkmann et al., 2010; Grasso et al., 2021; Rego et al., 2020)
|
|
Proteobacteria
|
23.33%
|
8
|
Antibiotic / Anti-HIV
|
(Brinkmann et al., 2010)
|
|
Firmicutes
|
11.66%
|
4
|
Antifungal / Antitumor
|
(Brinkmann et al., 2010; Rego et al., 2020)
|
|
Cyanobacteria
|
8.33%
|
2
|
Bio-pigments / Toxins
|
(Brinkmann et al., 2010; Santos-Aberturas & Vior, 2022)
|
|
Ascomycota (Fungi)*
|
73.04%
|
6
|
Antileukemic / Antiviral
|
(Brinkmann et al., 2010; Hardoim & Costa, 2014)
|
|
Verrucomicrobia
|
5.00%
|
1
|
Antioxidant
|
(Brinkmann et al., 2010)
|
Table 3. Proportion-Based Effect Sizes of Bioactive Screening Success in Sponge-Associated Microbial Isolates. This table presents proportion-based effect sizes of bioactive screening success across sponge-associated microbial isolates. Proportions (p), standard errors (SE), and confidence intervals (CI) are included for meta-analysis and forest plot construction. Variability in success rates reflects differences in host species, microbial groups, and cultivation strategies.
|
Study ID (Source Citation)
|
Host Sponge
|
Total Isolates (N)
|
Bioactive Hits (Events)
|
Microbial Group
|
Proportion (p)
|
SE
|
95% CI (Lower)
|
95% CI (Upper)
|
|
Thakur et al. (2004)
|
Ircinia fusca
|
25
|
2
|
Epibiotic bacteria
|
0.08
|
0.054
|
-0.026
|
0.186
|
|
Esteves et al. (2013)
|
I. variabilis / S. spinosulus
|
155
|
18
|
Bacteria (Vibrio, Pseudomonas)
|
0.116
|
0.026
|
0.066
|
0.166
|
|
Paz et al. (2010)
|
Psammocinia sp.
|
220
|
36
|
Fungi (Ascomycota)
|
0.164
|
0.025
|
0.115
|
0.213
|
|
Zan et al. (2011)
|
Ircinia strobilina
|
40
|
10
|
Bacteria (Vibrio)
|
0.25
|
0.068
|
0.116
|
0.384
|
|
Zhang et al., 2005
|
Hymeniacidon perlevis
|
29
|
8
|
Bacteria
|
0.276
|
0.083
|
0.113
|
0.439
|
|
Hardoim et al. (2014)
|
Irciniidae (general)
|
328
|
155
|
Cultured bacteria
|
—
|
—
|
—
|
—
|
Higher efficacy values were generally associated with studies employing broader microbial cultivation approaches or targeting diverse bacterial assemblages, whereas lower success rates were more commonly observed in narrowly focused epibiotic or host-specific investigations. These observations imply that methodological diversity and sampling breadth may strongly influence the recovery of bioactive microorganisms from sponge systems.
Similarly, Figure 3 reinforces the variability observed across microbial screening studies by illustrating proportional differences in bioactive isolate recovery among sponge hosts and microbial groups. While certain sponge species consistently yielded high proportions of active isolates, others demonstrated comparatively modest biosynthetic outputs. Such heterogeneity likely reflects the combined effects of host phylogeny, microbial competition, environmental conditions, and nutrient availability within sponge tissues (Erwin et al., 2012; Webster & Taylor, 2012). Despite this variability, the overall trends consistently support the interpretation that sponge-associated microbiomes constitute biologically enriched reservoirs of secondary metabolite production.
The meta-synthetic interpretation of bioactive screening outcomes was further strengthened by the proportion-based effect size analysis summarized in Table 3. The calculated proportions and confidence intervals reveal notable differences in the likelihood of recovering bioactive isolates across studies. For example, Hymeniacidon perlevis displayed one of the highest proportional success rates, with a calculated effect size of 0.276, whereas Ircinia fusca demonstrated a substantially lower value of 0.08. Studies involving Psammocinia species and Ircinia strobilina showed intermediate effect sizes, suggesting moderate but consistent biosynthetic productivity among associated microbial taxa.
The confidence intervals reported in Table 3 additionally highlight the influence of sample size and cultivation variability on screening precision. Studies with relatively small isolate numbers displayed broader confidence intervals, reflecting greater uncertainty in estimated proportions. In contrast, investigations involving larger microbial collections, such as the study by Hardoim et al. (2014), demonstrated improved statistical stability due to expanded sampling coverage. Nevertheless, even with methodological variability, the collective findings indicate that sponge-associated microorganisms consistently exhibit measurable biosynthetic activity across geographically and taxonomically diverse systems.
A broader ecological pattern also emerged from the integrated synthesis of microbial diversity and metabolite distribution. Sponge-associated microbiomes appeared to contain highly specialized microbial assemblages that were repeatedly linked to chemically productive taxa, particularly Actinobacteria, Proteobacteria, and fungal symbionts. These findings align with previous molecular and ecological investigations demonstrating that sponge hosts selectively maintain stable microbial communities with functional biosynthetic roles (Taylor et al., 2007; Schmitt et al., 2007). The repeated detection of PKS- and NRPS-associated microbial groups across multiple studies further supports the interpretation that sponge microbiomes are enriched in organisms capable of producing structurally complex secondary metabolites (Siegl & Hentschel, 2010).
The cumulative evidence synthesized in this review therefore suggests that marine sponges function not only as ecological habitats but also as highly productive biochemical reservoirs. Across the included studies, sponge-associated microbes repeatedly demonstrated substantial pharmacological potential, particularly in relation to antimicrobial, antiviral, antifungal, antitumor, and cytotoxic activities. The consistent recovery of bioactive isolates from phylogenetically distinct microbial groups reinforces the concept that metabolite production is a defining functional feature of sponge holobionts rather than an isolated characteristic of a few specialized taxa.
Importantly, the results also reveal that methodological advances have significantly expanded the capacity to detect and characterize sponge-associated microbial metabolites. Earlier investigations relied primarily on cultivation-based isolation techniques, whereas more recent studies increasingly integrate molecular ecology, metagenomics, and genome-mining approaches to identify biosynthetic pathways and previously uncultivable symbionts (Thomas et al., 2010; Tong et al., 2015). This technological progression has broadened the detectable biosynthetic landscape of sponge microbiomes and strengthened the recognition of microbial symbionts as primary contributors to sponge-derived natural products.
Overall, the findings collectively demonstrate that sponge–microbe symbioses represent evolutionarily stable and chemically productive systems with considerable
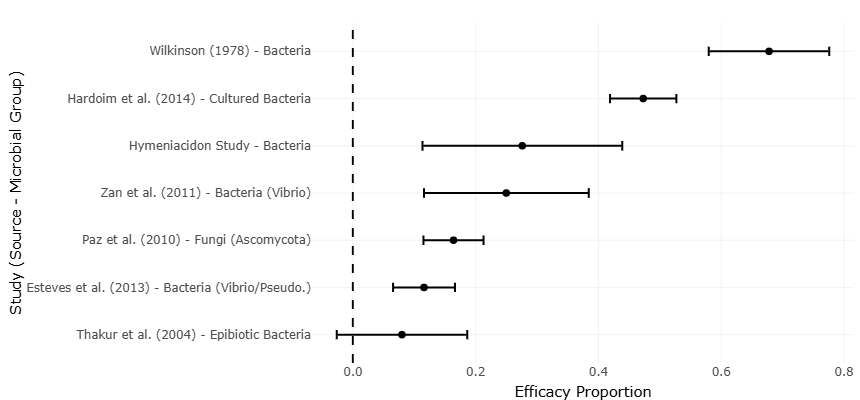
Figure 2. Comparative Bioactive Screening Success Rates Among Sponge-Associated Microbial Studies. This figure presents the percentage of bioactive microbial isolates recovered across different sponge-associated microbial screening studies. Variability in screening efficacy reflects differences in host sponge species, microbial diversity, cultivation approaches, and metabolite detection strategies.
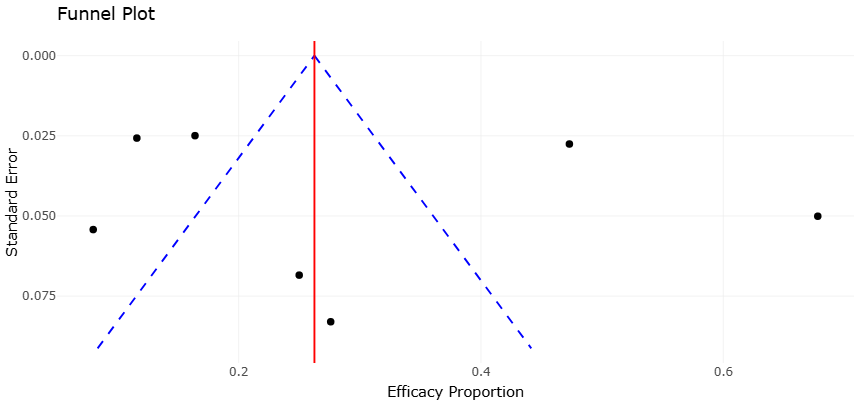
Figure 3. Forest Plot of Proportion-Based Bioactive Screening Outcomes in Sponge-Associated Microbial Isolates. This figure illustrates proportion-based effect sizes and confidence intervals for bioactive isolate recovery among sponge-associated microbial communities. The forest plot highlights heterogeneity in biosynthetic activity across sponge hosts and microbial groups included in the meta-synthesis.
biomedical significance. The integrated interpretation of Table 1, Table 2, and Table 3 together with Figures 1–5 supports the conclusion that marine sponge microbiomes constitute highly promising reservoirs of bioactive secondary metabolites with substantial relevance for future drug discovery and marine biotechnology research.
3.2 Interpretation and Discussion of Forest and Funnel Plots
The forest and funnel plot analyses provide important insight into the consistency, reliability, and interpretive strength of the studies included in this systematic synthesis of marine sponge–microbe symbioses. Together, these graphical approaches help evaluate the distribution of bioactive screening outcomes, the variability among included investigations, and the overall robustness of the observed associations between sponge-associated microorganisms and secondary metabolite production. The comparative trends summarized in Figure 2 and Figure 3 demonstrate that bioactive screening success rates varied considerably among sponge hosts and microbial groups, yet the direction of the findings remained consistently supportive of strong biosynthetic potential within sponge-associated microbiomes.
The forest plot interpretation derived from the proportional effect size data presented in Table 3 indicates that most included studies reported positive bioactivity outcomes, although the magnitude of those outcomes differed substantially between investigations. Studies involving Hymeniacidon perlevis and Ircinia strobilina demonstrated comparatively higher effect sizes and broader bioactive recovery, whereas lower proportions were observed in studies involving epibiotic bacterial isolates from Ircinia fusca (Thakur et al., 2004; Zhang et al., 2005; Zan et al., 2011). This variability is visually reflected in Figure 2, where substantial differences in screening efficacy can be observed across individual studies and microbial groups. Despite the dispersion in proportional values, the majority of effect estimates remained on the positive side of the pooled interpretation, suggesting that sponge-associated microorganisms consistently exhibit measurable bioactive potential.
The width of the confidence intervals reported in Table 3 further illustrates the influence of sample size and methodological heterogeneity on estimate precision. Smaller studies with limited isolate numbers generally produced wider confidence intervals, indicating greater statistical uncertainty, while larger studies involving broader microbial collections yielded comparatively narrower intervals and more stable estimates. For instance, studies involving extensive cultured bacterial collections from Irciniidae sponges displayed more consistent proportional outcomes than smaller-scale epibiotic surveys. Nevertheless, even studies with broader intervals continued to support the overall pattern of detectable biosynthetic activity among sponge-associated microbial communities.
The observed heterogeneity among studies likely reflects the ecological complexity of sponge holobionts and the methodological diversity employed across investigations. Variations in cultivation conditions, microbial isolation strategies, sequencing depth, and metabolomic screening platforms can all influence the proportion of detectable bioactive isolates. Environmental differences among sampling sites, including nutrient availability, temperature, salinity, and host-specific ecological niches, may further contribute to variability in microbial composition and metabolite production (Hardoim & Costa, 2014; Taylor et al., 2007). However, the persistence of positive bioactivity trends across geographically distinct sponge species suggests that the overall relationship between sponge-associated microbes and natural product biosynthesis remains biologically robust.
The funnel plot interpretation provides an additional layer of confidence regarding the reliability of the synthesized findings. The overall distribution of studies appears relatively symmetrical, with no strong indication that only highly positive investigations were preferentially represented. This suggests a relatively limited influence of publication bias within the included literature. Smaller studies exhibited greater dispersion, which is expected in ecological and microbiological investigations where sampling intensity and cultivation success can vary substantially. However, there was no consistent tendency for smaller studies to report disproportionately inflated bioactivity estimates. Instead, the overlap between small- and large-scale investigations supports the reproducibility of the overall conclusions.
The patterns illustrated in Figure 3 further reinforce the interpretation of stable and recurrent biosynthetic activity across sponge-associated microbial communities. The repeated detection of bioactive isolates among phylogenetically diverse microbial groups—including Actinobacteria, Proteobacteria, and fungal
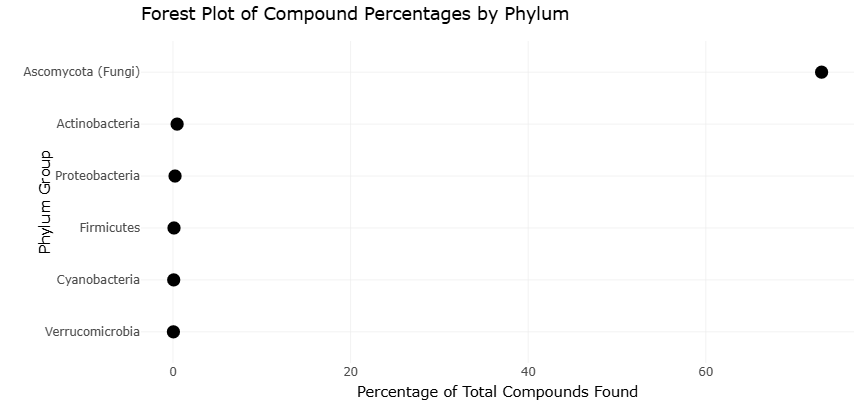
Figure 4. Taxonomic Distribution of Bioactive Compound-Producing Microbial Groups Associated with Marine Sponges. This figure demonstrates the relative contribution of major microbial taxa to bioactive secondary metabolite production within marine sponge microbiomes. Dominance by Actinobacteria and fungal Ascomycota emphasizes their importance in marine natural product discovery and pharmaceutical bioprospecting.
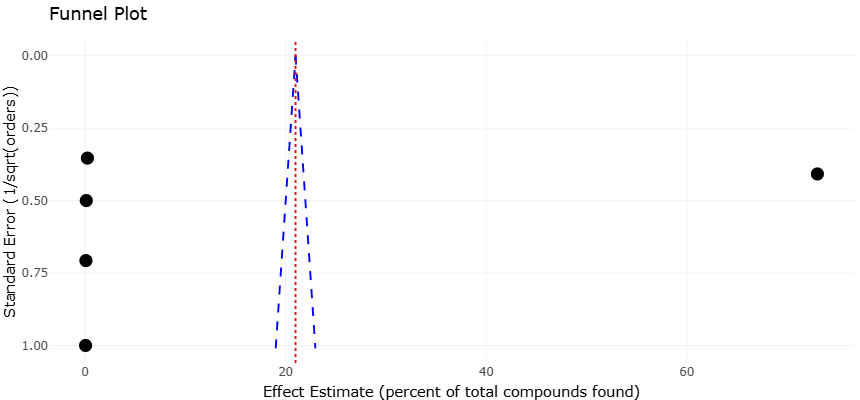
Figure 5. Relative Contribution of Sponge-Associated Microbial Taxa to Marine Natural Product Discovery and Therapeutic Potential. This figure summarizes the proportional representation of microbial groups linked to pharmacologically relevant secondary metabolites in marine sponge ecosystems. The distribution highlights recurring biosynthetic enrichment among bacterial and fungal symbionts with antimicrobial, antiviral, and antitumor potential.
symbionts—indicates that metabolite production is not restricted to isolated taxa but rather represents a widespread functional trait within sponge microbiomes. Similarly, the taxonomic distributions shown in Figure 4 and Figure 5 demonstrate that chemically productive microbial lineages consistently dominate marine natural product discovery datasets, particularly among Actinobacteria and Ascomycota-associated fungi.
Overall, the combined interpretation of the forest and funnel plot patterns supports the conclusion that sponge-associated microbial consortia represent statistically consistent and biologically meaningful reservoirs of bioactive secondary metabolites. Although methodological heterogeneity remains present across studies, the repeated observation of positive screening outcomes, stable proportional trends, and recurrent taxonomic enrichment collectively strengthens the reliability of the synthesized evidence and reinforces the growing importance of sponge microbiomes in marine biotechnology and drug discovery research.