1. Introduction
The rapid intensification of global livestock production has become one of the defining agricultural transitions of the twenty first century. As the global population moves steadily toward an estimated 9.7 billion by 2050, pressure on food systems continues to mount in ways that are increasingly difficult to ignore. Demand for meat, milk, and other animal derived products is rising not only because of population growth, but also due to urbanization, dietary transitions, and expanding middle income populations across developing economies (Alexandratos & Bruinsma, 2012; Keating et al., 2014). Current projections suggest that global livestock production may need to increase by nearly 70% over coming decades to satisfy anticipated consumption patterns (Hunter et al., 2017). Yet this expansion introduces a persistent and uncomfortable question: can livestock systems continue to grow without simultaneously deepening environmental instability, resource depletion, and food insecurity?
Modern animal feeding systems remain heavily dependent on conventional feed commodities, particularly cereals and soybean meal. Vast quantities of grains that might otherwise contribute directly to human nutrition are instead redirected into livestock production chains. Estimates indicate that nearly 900 million tons of cereals and a substantial majority of globally produced soybean meal are used annually in animal feed formulations (Henchion et al., 2017; Nasseri et al., 2011). While this system has undeniably supported remarkable productivity gains, it has also intensified the long standing “food versus feed” debate, especially in regions where nutritional insecurity persists. Soybean expansion, meanwhile, continues to reshape landscapes on a massive scale. In countries such as Brazil, soybean cultivation has been strongly associated with deforestation, biodiversity loss, and ecological fragmentation within sensitive ecosystems including the Amazon and Cerrado regions (Lima et al., 2019). These environmental pressures are compounded further by greenhouse gas emissions and soil degradation linked to large scale feed crop production (Rojas Downing et al., 2017).
Ruminant agriculture occupies a particularly complicated position within this sustainability discussion. On one hand, ruminants possess the remarkable physiological ability to convert fibrous plant biomass that humans cannot digest into nutrient dense food products. This capability represents an important biological advantage within global food systems (Annison & Bryden, 1998; National Research Council, 2001). On the other hand, ruminant production systems are also major contributors to agricultural methane emissions, which significantly influence the global climate burden (Rojas Downing et al., 2017). The efficiency of ruminal fermentation is shaped by an intricate interplay among diet composition, microbial ecology, feed additives, and host metabolism. Consequently, nutritional strategies capable of improving feed efficiency while simultaneously moderating methane production have attracted growing scientific and commercial attention.
Against this backdrop, alternative protein sources and functional feed ingredients are increasingly being explored as part of more sustainable livestock production models. Among these, single cell proteins (SCPs) have emerged as particularly promising candidates. SCPs are microbial biomasses derived from bacteria, algae, yeasts, or fungi that contain high concentrations of protein and other valuable nutrients (Bratosin et al., 2021; Nasseri et al., 2011). Interest in SCP technology is not entirely new; however, recent concerns surrounding land use, feed security, and circular bioeconomy frameworks have renewed enthusiasm for microbial based feed production systems. Rather than relying exclusively on conventional agricultural inputs, SCPs may allow the conversion of industrial by products and organic residues into nutritionally valuable biomass.
Within this broader category, Aspergillus oryzae has received increasing attention due to its unique combination of safety, metabolic versatility, and long standing industrial relevance. Historically, A. oryzae has been used extensively in East Asian food fermentations, particularly in the production of soy sauce, sake, and miso (Bentley, 2006). Unlike many microorganisms that require tightly controlled sterile conditions, A. oryzae demonstrates considerable adaptability across diverse substrates and environmental conditions. Its extracellular enzyme systems—including proteases, amylases, cellulases, and lipases—allow efficient degradation of complex carbohydrates and organic materials (Jin et al., 2001; Ferreira et al., 2016). This enzymatic flexibility has positioned the fungus as a potential cornerstone organism in waste valorization systems.
Several studies have demonstrated the feasibility of cultivating A. oryzae on industrial side streams and agro industrial waste products. Thin stillage generated during ethanol production, sugarcane vinasse, and food processing wastewaters have all been investigated as potential substrates for fungal biomass generation (Duru & Uma, 2003; Ferreira et al., 2014; Ferreira et al., 2016). In many respects, this approach aligns closely with circular economy principles, where low value waste streams are redirected into productive nutritional applications rather than discarded. Yet despite the conceptual attractiveness of such systems, questions surrounding scalability, contamination control, and economic viability remain only partially resolved.
Nutritionally, A. oryzae biomass appears highly competitive with several conventional protein ingredients. Reported crude protein concentrations commonly range between 40% and 60% of dry matter, accompanied by appreciable levels of essential amino acids such as lysine and threonine (Karimi et al., 2021). Although sulfur containing amino acids may occasionally represent a limiting factor, the overall amino acid balance compares favorably with soybean meal and, in some contexts, even fishmeal (Karimi et al., 2021; Nasseri et al., 2011). Additionally, fungal biomass contains bioactive lipids, minerals, and micronutrients that may contribute functional nutritional benefits beyond simple protein replacement.
Importantly, the role of A. oryzae in ruminant feeding systems extends beyond its nutritional composition alone. A substantial body of research has explored its function as a direct fed microbial or fermentation extract capable of influencing ruminal microbial dynamics. Early investigations reported that A. oryzae fermentation products could stimulate fibrolytic bacterial populations and enhance degradation of neutral detergent fiber and acid detergent fiber within the rumen (Beharka & Nagaraja, 1993; Beharka & Nagaraja, 1998). Such microbial shifts appear to promote greater production of volatile fatty acids, particularly acetate and propionate, which are central energy substrates for ruminants (Beharka & Nagaraja, 1998).
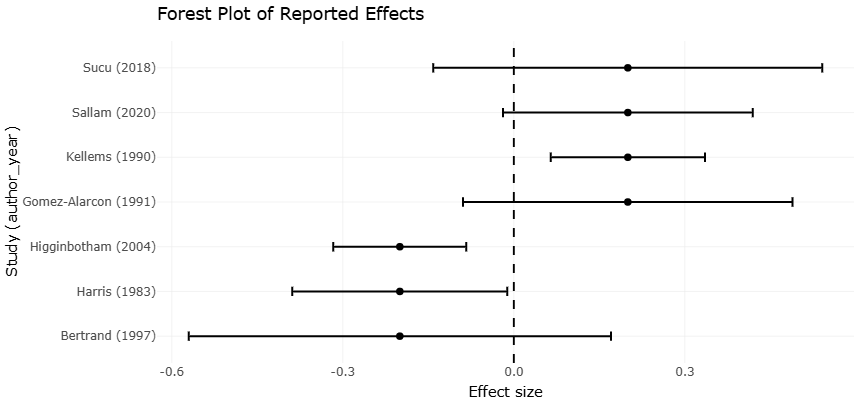
The mechanisms involved are likely multifactorial and perhaps not yet fully understood. Some evidence suggests that A. oryzae supplementation may stabilize ruminal pH through support of lactate utilizing bacteria such as Selenomonas ruminantium and Megasphaera elsdenii. This effect may be particularly valuable in high concentrate feeding systems where rapid carbohydrate fermentation can predispose animals to subacute ruminal acidosis and impaired productivity. Improvements in feed digestibility, dry matter intake, and milk production have been documented across multiple dairy and beef production studies (Bertrand & Grimes, 1997; Chiou et al., 2002; Chiquette, 1995; Gomez Alarcon et al., 1991; Gomez-Alarcon et al., 1991; Kellems et al., 1990; Sallam et al., 2020; Sallam et al., 2020; Sucu et al., 2018; Takiya et al., 2017).
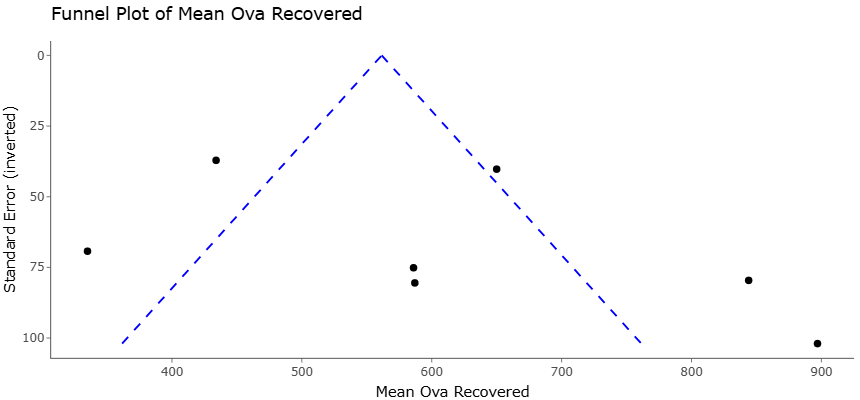
Nevertheless, the literature remains far from entirely consistent. Certain trials have reported only marginal benefits, while others observed negligible or variable responses depending on diet composition, supplementation dose, stage of lactation, or animal physiological condition (Harris et al., 1983; Higginbotham et al., 2004; Higginbotham et al., 2004). This inconsistency may reflect the inherent complexity of ruminal ecosystems, where microbial interactions are influenced by numerous nutritional and environmental variables simultaneously. It also highlights an important limitation within the current evidence base: many feeding studies remain relatively small, context specific, or methodologically heterogeneous.
Consequently, systematic reviews and meta analytic approaches have become increasingly necessary to clarify broader patterns across independent studies. By synthesizing data across production systems and dietary conditions, meta analyses may improve precision in estimating the effects of A. oryzae supplementation while also identifying important moderators of response. Such analyses are particularly relevant for disentangling whether observed benefits arise primarily from enhanced fiber digestion, altered microbial fermentation pathways, improved nutrient utilization, or indirect metabolic effects.
Beyond productivity considerations, A. oryzae may also contribute to environmental mitigation strategies within livestock systems. Some emerging evidence suggests that redirecting ruminal hydrogen flow toward propionate formation rather than methanogenesis could reduce methane emissions intensity (Choudhury et al., 2022). If consistently validated, such effects would position fungal based feed technologies within broader climate adaptation and mitigation frameworks for agriculture.
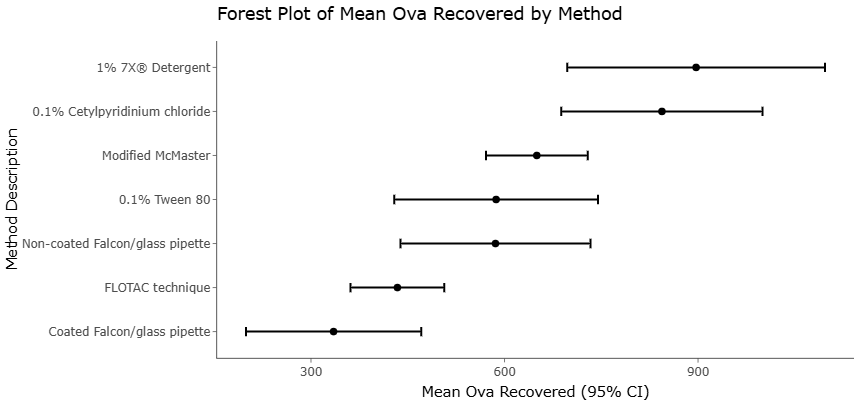
Still, optimism surrounding fungal biomass technologies should probably be tempered with caution. The use of wastewater-derived substrates and organic residues inevitably raises concerns about pathogen contamination and biosafety. Detection and monitoring methodologies for soil transmitted helminths and related contaminants therefore remain critically important within fungal biomass production systems (Amoah et al., 2017; Jeandron et al., 2014; Ravindran et al., 2019; Ravindran et al., 2019). Regulatory oversight, quality assurance protocols, and economic feasibility analyses will likely determine whether these technologies can transition from experimental promise to commercially viable implementation.
Ultimately, the growing interest in Aspergillus oryzae reflects a broader shift occurring within livestock nutrition science itself. Feed systems are no longer evaluated solely through the lens of productivity. Increasingly, they are being examined according to their environmental efficiency, resilience, waste recovery potential, and compatibility with sustainable food system goals. Within this evolving landscape, A. oryzae represents not merely a microbial feed additive, but a potentially important intersection between microbial biotechnology, circular bioeconomy strategies, and sustainable ruminant agriculture.