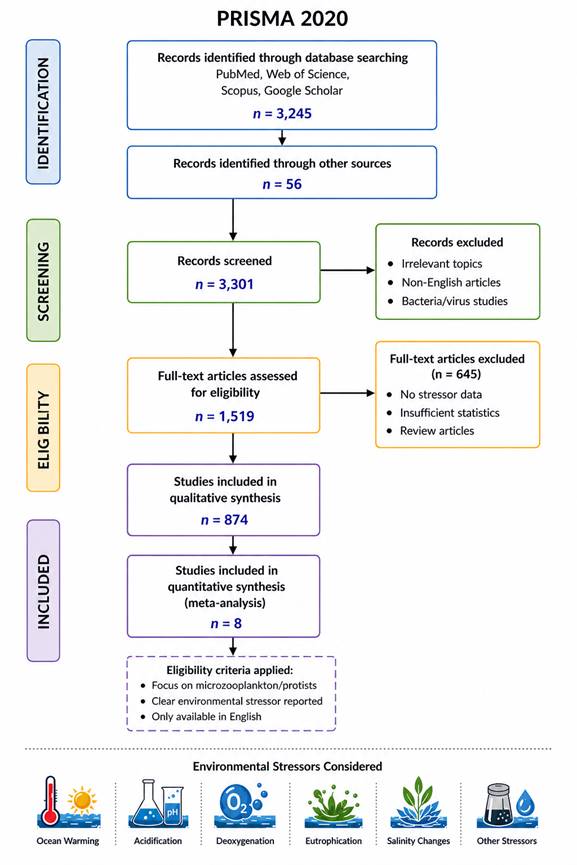
1.Introduction
Microzooplankton and protist communities form the living engine of marine food webs, quietly governing the flow of energy and matter across the ocean. Although often overlooked because of their microscopic size, these organisms represent an extraordinary share of marine eukaryotic diversity and play indispensable roles as primary consumers, secondary producers, decomposers, and recyclers of nutrients (Fenchel, 2008; De Vargas et al., 2015). Through intense grazing pressure on phytoplankton and bacteria, microzooplankton process an estimated 20–30 Pg of carbon annually—more than twice the amount consumed by mesozooplankton—placing them at the center of global carbon cycling and climate regulation (Buitenhuis et al., 2010; Worden et al., 2015).
Despite their centrality to ecosystem functioning, microzooplankton communities are increasingly exposed to multiple, overlapping global stressors driven by climate change and human activity. Ocean warming, acidification, deoxygenation, eutrophication, and altered nutrient stoichiometry are reshaping marine habitats at unprecedented rates (Bindoff et al., 2019). Understanding how these pressures influence microzooplankton vulnerability, resilience, and adaptive capacity has therefore become a critical challenge for marine ecology. Recent synthesis efforts emphasize that ecosystem risk cannot be inferred solely from exposure; rather, it emerges from the interaction between exposure, biological sensitivity, and adaptive capacity at both species and community levels (López-Abbate, 2021).
This introduction draws on a systematic review and meta-analytical interpretation of peer-reviewed studies to examine how global environmental hazards affect microzooplankton communities, while highlighting the biological traits and community processes that buffer ecosystem functioning. By integrating quantitative effect sizes, experimental evidence, and network-based analyses, the review frames microzooplankton not simply as vulnerable components of marine ecosystems, but as dynamic and resilient systems capable of reorganization under stress.
The foundational role of microzooplankton is rooted in the microbial loop, a conceptual framework that recognizes microbes as dominant agents of carbon and nutrient recycling in the sea (Fenchel, 2008). By grazing on picophytoplankton and heterotrophic bacteria that are inaccessible to larger consumers, microzooplankton repackage dissolved and particulate organic matter into forms that can move upward through food webs or sink to depth (Buitenhuis et al., 2010; Worden et al., 2015). This function becomes especially important in oligotrophic oceans, where smaller cells dominate primary production and classical food chains are truncated.
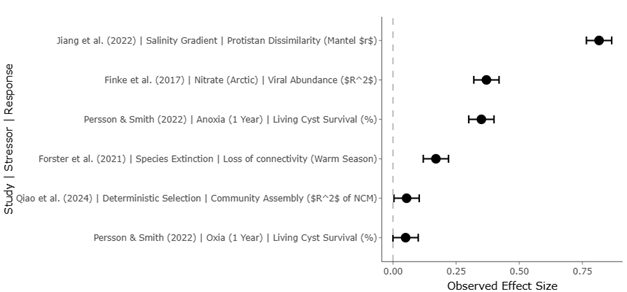
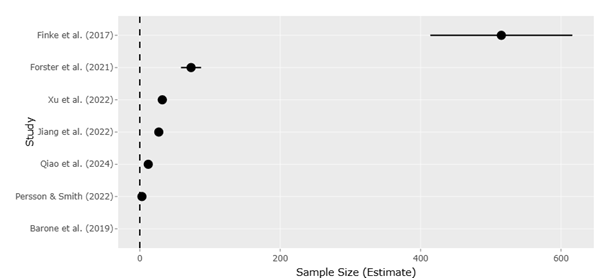
Beyond carbon transfer, microzooplankton influence nutrient regeneration, trace gas production, and microbial diversity through selective grazing (Mitra et al., 2014; Park et al., 2014). Viral interactions further complicate these dynamics, as viral infection of microbial hosts redirects organic matter through the viral shunt, enhancing nutrient recycling while limiting carbon export (Wilhelm & Suttle, 1999; Suttle, 2007). Meta-analytical evidence indicates that viral abundance is strongly linked to environmental drivers such as nitrogen and phosphorus availability, highlighting the sensitivity of microbial mortality pathways to nutrient imbalance (Finke et al., 2017).
Among climate-related stressors, ocean warming represents one of the most pervasive and rapidly intensifying hazards. Temperature exerts direct control over metabolic rates, growth, and grazing activity in heterotrophic protists (Rose & Caron, 2007). Evidence synthesized across latitudinal gradients suggests that warming generally favors smaller, fast-growing protists, promoting poleward range expansions and seasonal shifts in community composition (Edwards et al., 2006; Jonkers et al., 2019). While polar and subpolar ecosystems may experience amplified responses due to historical thermal constraints, many taxa demonstrate high adaptive capacity by tracking shifting thermal niches and prey phenology (Field et al., 2006).
Ocean acidification (OA), by contrast, is characterized by high exposure but comparatively low sensitivity for most non-calcifying microzooplankton (Nielsen et al., 2010; Suffrian et al., 2008). Experimental and field studies indicate that projected end-of-century pH changes exert limited direct effects on growth and grazing for many taxa. Instead, community-level impacts are often indirect, mediated through changes in phytoplankton food quality and elemental composition (Park et al., 2014). Calcifying organisms such as planktonic foraminifera remain an exception, exhibiting measurable reductions in shell weight and calcification under acidified conditions (Moy et al., 2009).
In contrast to warming and acidification, deoxygenation emerges as the hazard associated with the highest ecological risk. Declining oxygen concentrations directly constrain aerobic metabolism, reduce grazing rates, and restructure species assemblages (Breitburg et al., 2018). Ciliates, in particular, exhibit narrow oxygen niches and pronounced reductions in growth under hypoxic conditions (Rocke & Liu, 2014). Meta-analytical evidence further demonstrates that deoxygenation drives strong species replacement and diversity loss, increasing community vulnerability despite the persistence of overall function (López-Abbate, 2021).
In coastal and estuarine systems, anthropogenic eutrophication interacts with natural gradients such as salinity to shape microzooplankton assemblages. Elevated nutrient inputs often weaken trophic coupling by inducing feeding saturation, whereby grazers cannot control rapidly increasing prey biomass (Redden et al., 2002; Heisler et al., 2008). This decoupling favors the proliferation of unpalatable or harmful algal species, many of which rely on mixotrophy as a dominant nutritional strategy (Burkholder et al., 2008; Mitra et al., 2014).
Salinity gradients impose additional constraints, acting as strong environmental filters that limit dispersal and select for specialized taxa. Large-scale sequencing studies reveal that salinity explains a substantial proportion of community dissimilarity in estuarine protists, often exceeding the influence of temperature or nutrients (Jiang et al., 2022; Xu et al., 2022). These findings underscore the importance of regional context in assessing ecological risk, particularly in highly modified coastal ecosystems.
A defining feature of microzooplankton communities is their extraordinary adaptive capacity, rooted in functional diversity, rapid generation times, and access to a vast “rare biosphere” of low-abundance taxa (López-Abbate, 2021). Mixotrophy exemplifies this flexibility, allowing organisms to combine phototrophy and phagotrophy to exploit fluctuating resource landscapes (Mitra et al., 2014). Stoichiometric plasticity further enables protists to adjust internal nutrient ratios, sustaining growth even when prey quality declines (Mitra et al., 2014).
Life-cycle strategies such as resting cyst formation provide additional resilience. Under anoxic conditions, dinoflagellate cysts can remain viable for decades or even centuries, effectively functioning as biological seed banks that reintroduce diversity following disturbance (Lundholm et al., 2011; Persson & Smith, 2022). While this mechanism enhances long-term persistence, it can also amplify ecological risk by facilitating recurrent harmful algal blooms when favorable conditions return.
The limits of microzooplankton resilience are perhaps best illustrated by their presence in extreme habitats such as Deep-sea Hypersaline Anoxic Basins (DHABs). In these environments, marine fungi and other protists thrive under extreme salinity, toxicity, and hydrostatic pressure, actively contributing to organic matter remineralization (Gostinčar et al., 2011; Barone et al., 2019). Such systems highlight the remarkable physiological and metabolic plasticity of micro-eukaryotes, reinforcing their role as ubiquitous and adaptable components of marine ecosystems.
Advances in high-throughput sequencing and bioinformatics have further transformed understanding of community resilience. Amplicon sequence variant (ASV)-based approaches, such as DADA2, provide fine-scale resolution of protistan diversity, allowing robust detection of subtle shifts in community structure (Callahan et al., 2016). Network analyses reveal that climate stress tends to reduce interaction complexity and increase vulnerability to species loss, as demonstrated in long-term lake studies where warming disrupted traditional plankton succession (Forster et al., 2021). Yet even in simplified networks, functional roles are often maintained through species replacement, underscoring the stabilizing influence of redundancy.
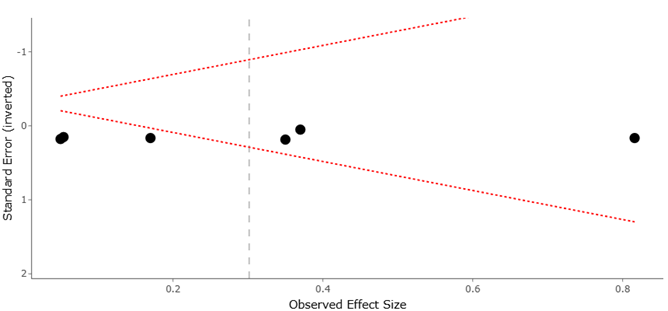
Collectively, evidence synthesized from systematic review and meta-analysis supports a nuanced view of microzooplankton vulnerability. While individual taxa may exhibit high sensitivity to specific stressors—particularly deoxygenation—the community as a whole often maintains functional stability through adaptive traits and species replacement (López-Abbate, 2021; Qiao et al., 2024). This balance between sensitivity and resilience positions microzooplankton as both indicators of environmental change and key buffers against ecosystem collapse. Understanding their responses is therefore essential for predicting the future trajectory of marine biogeochemical cycles in an increasingly stressed ocean.