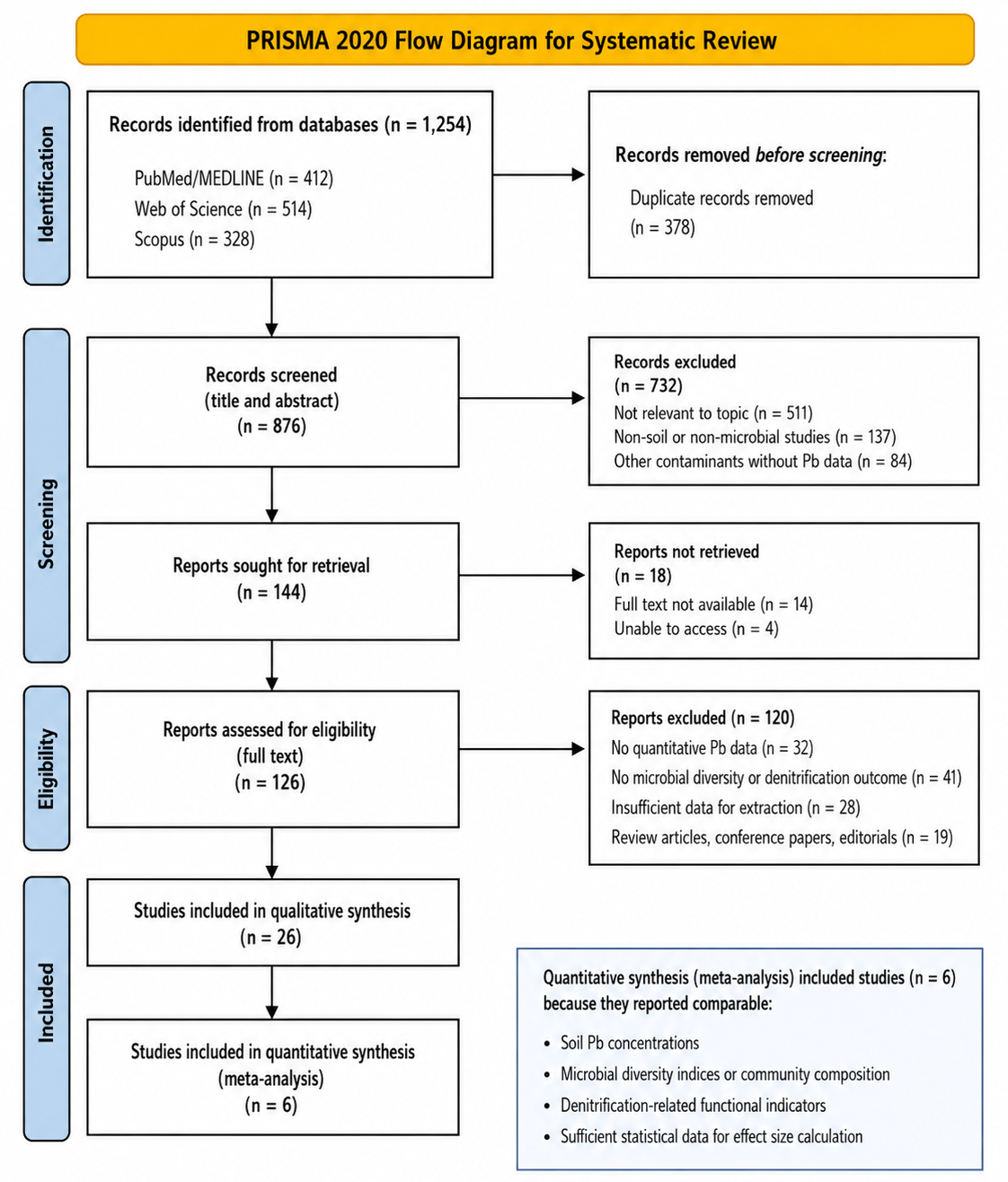
1. Introduction
Lead (Pb) contamination has become one of the more persistent and ecologically disruptive forms of soil pollution associated with industrialization, mining, smelting activities, urban expansion, and intensive agricultural practices. Although soils are often perceived merely as physical substrates that support plant growth, they are, in reality, extraordinarily dynamic biological systems inhabited by diverse microbial communities that regulate nutrient cycling, organic matter decomposition, carbon sequestration, and ecosystem productivity (Fierer, 2017). The integrity of these microbial networks is deeply intertwined with soil health and ecological resilience, meaning that disturbances affecting microbial diversity may ultimately compromise broader ecosystem functioning (Delgado-Baquerizo et al., 2016). In recent years, growing concern has emerged over how toxic metals, particularly Pb, alter these belowground biological systems in ways that are not always immediately visible but may have long-lasting environmental consequences.
Unlike certain trace metals that serve physiological functions at low concentrations, Pb has no known biological role in microbial metabolism and is considered toxic even at relatively low exposure levels (Violante et al., 2010). What makes Pb especially problematic is not merely its toxicity, but its persistence. Once introduced into soils, Pb strongly associates with clay particles, oxides, and organic matter, allowing it to remain in the environment for decades or even centuries (Alengebawy et al., 2021). This prolonged residence time means that microbial communities are often exposed to chronic rather than acute contamination, creating sustained ecological pressure that may reshape community composition, reduce biodiversity, and alter functional processes over time (Giller et al., 2009). In heavily contaminated environments such as mining districts and industrial zones, these changes can become profound enough to transform the biological identity of the soil itself.
Yet, the ecological response to Pb contamination is rarely straightforward. Soil microbial communities do not respond uniformly across landscapes or contamination gradients. Instead, responses appear to depend heavily on factors such as pH, organic matter content, moisture, redox potential, metal speciation, and interactions with co-occurring contaminants (Zhao et al., 2020). Acidic soils, for instance, often increase Pb mobility and bioavailability, intensifying microbial exposure and toxicity, whereas soils rich in organic matter may partially immobilize Pb through chelation and adsorption processes (Violante et al., 2010). This environmental complexity makes it difficult to predict microbial outcomes using Pb concentration alone. In some soils, modest contamination may produce little detectable effect, while in others, similar concentrations may trigger sharp reductions in microbial diversity and metabolic activity.
Over the past decade, advances in molecular ecology have significantly improved understanding of how microbial communities respond to heavy metal stress. Traditional cultivation-based approaches captured only a fraction of microbial diversity, often overlooking rare or unculturable taxa that nonetheless contribute meaningfully to ecosystem stability. More recently, tools such as PCR-DGGE, high-throughput 16S rRNA sequencing, metagenomics, and functional gene analyses have enabled researchers to investigate microbial assemblages with far greater resolution (Bhakta et al., 2018). These approaches have revealed that Pb contamination can reshape both taxonomic composition and ecological interactions within soil microbiomes. Certain microbial groups decline rapidly under metal stress, whereas others persist or even proliferate, suggesting varying levels of resistance and adaptive capacity among taxa (Lin et al., 2019).
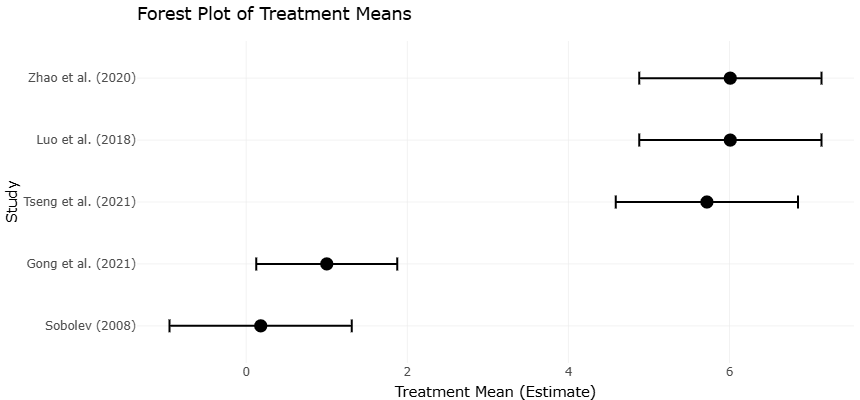
A recurring observation across contaminated environments is the decline in microbial diversity and richness as heavy metal burdens increase. Studies conducted near smelting sites, mining areas, and contaminated farmlands consistently report reductions in indices such as Shannon diversity and Chao1 richness under elevated Pb concentrations (Luo et al., 2018; Tseng et al., 2021; Zhao et al., 2020). These declines likely reflect selective pressures favoring metal-tolerant microorganisms while sensitive taxa diminish or disappear. Although some degree of community restructuring may represent ecological adaptation, persistent biodiversity loss raises concerns regarding ecosystem multifunctionality because diverse microbial communities tend to support more stable and efficient biogeochemical processes (Delgado-Baquerizo et al., 2016).
Interestingly, the relationship between Pb contamination and microbial diversity does not always appear linear. Several investigations suggest the existence of threshold-dependent responses in which microbial communities remain relatively stable up to a certain contamination level before experiencing abrupt structural collapse (Lin et al., 2019; Gao et al., 2019). Such ecological thresholds are particularly important because they imply that microbial systems may tolerate gradual environmental stress only to a point, beyond which resilience mechanisms become overwhelmed. This notion aligns with broader ecological theories concerning tipping points and nonlinear ecosystem responses under persistent disturbance. Understanding where these thresholds occur may therefore be essential for environmental risk assessment and remediation planning.
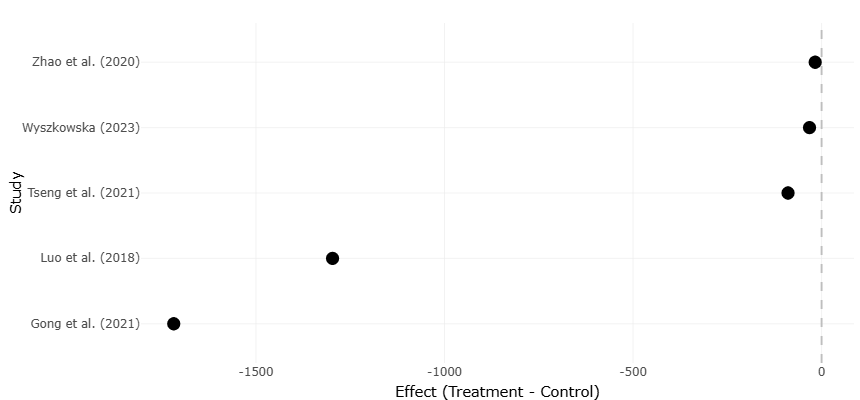
Beyond shifts in diversity, Pb contamination also influences microbial functionality, particularly processes associated with nitrogen cycling. Denitrification, the stepwise microbial reduction of nitrate to gaseous nitrogen compounds, is especially sensitive to heavy metal stress because many of the enzymes involved are membrane-bound or periplasmic and thus more exposed to environmental contaminants (Liu et al., 2018; Sobolev & Begonia, 2008). Disruption of denitrifying communities may reduce nitrogen transformation efficiency, increase nitrate accumulation, and contribute to elevated emissions of nitrous oxide (N₂O), a potent greenhouse gas. Functional disturbances of this kind suggest that Pb contamination affects not only “who is present” within microbial communities but also “what these communities are capable of doing.” In some contaminated soils, reductions in microbial biomass, respiration, and enzyme activity have been observed alongside compositional changes, reinforcing the idea that structural and functional responses are closely linked (Hu et al., 2019; Fajardo et al., 2019; Gong et al., 2021).
At the same time, microbial communities possess varying degrees of resistance, resilience, and redundancy that may buffer ecosystems against environmental disturbance. Resistance refers to the ability of microbial communities to remain relatively unchanged despite stress, whereas resilience describes the capacity to recover after disturbance (Allison & Martiny, 2008; Shade et al., 2012). Functional redundancy further suggests that multiple microbial taxa may perform similar ecological roles, allowing critical processes to continue even when some species decline (Griffiths & Philippot, 2012). However, the extent to which redundancy truly protects ecosystem functioning under chronic Pb exposure remains uncertain. Rare microbial taxa, often overlooked because of their low abundance, may play disproportionately important ecological roles during environmental stress or recovery phases (Jousset et al., 2017). Consequently, even subtle diversity losses may have implications that are not immediately apparent.
Another complicating factor is the temporal dimension of contamination. Microbial communities in long-term contaminated soils may differ substantially from those exposed to recent Pb inputs. Chronic contamination can drive ecological adaptation, leading to communities dominated by resistant taxa that tolerate heavy metal stress more effectively (Beattie et al., 2018). While such adaptation may preserve some ecosystem functions, it can also fundamentally alter microbial interactions, metabolic pathways, and nutrient dynamics. Moreover, adapted communities may not readily revert to their original states following remediation efforts, suggesting that historical contamination can leave lasting ecological legacies (Kuppusamy et al., 2016).
The effects of Pb are also rarely isolated. Heavy metal-contaminated soils often contain mixtures of pollutants, including cadmium (Cd), zinc (Zn), chromium (Cr), copper (Cu), pesticides, and metalloids, which may interact synergistically or antagonistically (Alengebawy et al., 2021; Wyszkowska et al., 2023). These interactions complicate interpretation because microbial responses observed in field studies may reflect combined contaminant pressures rather than Pb alone. For example, co-contaminants may intensify oxidative stress, alter microbial membrane integrity, or shift competitive interactions within microbial networks (Cui et al., 2018; Gao et al., 2019). Consequently, understanding Pb toxicity requires a more integrative perspective that considers the broader physicochemical context of contaminated soils.
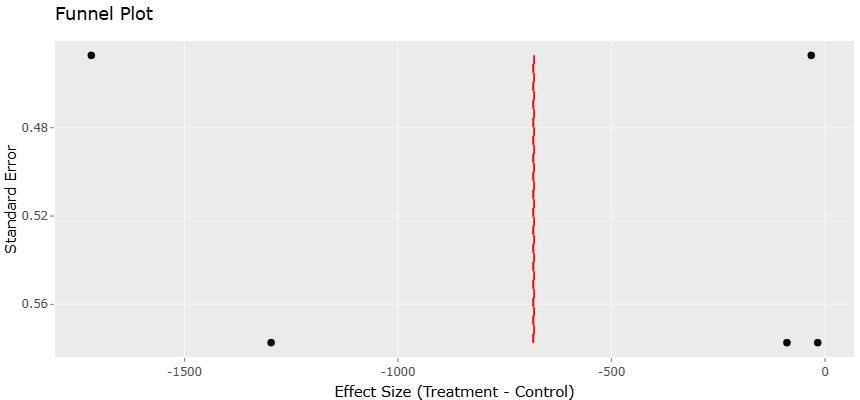
Despite substantial research activity, findings across individual studies remain fragmented and sometimes inconsistent due to differences in soil characteristics, contamination histories, molecular methods, and ecological metrics. Systematic reviews and meta-analytical approaches therefore provide an important opportunity to synthesize available evidence, identify broader ecological patterns, and evaluate whether threshold-dependent responses consistently emerge across studies. Meta-analysis also allows for the assessment of publication bias and variability among datasets through approaches such as funnel plot analysis and asymmetry testing (Egger et al., 1997; Sterne et al., 2011). Such synthesis is increasingly necessary as environmental scientists seek not only to describe contamination effects, but also to predict ecological outcomes and guide remediation strategies more effectively.
In this context, the present systematic review examines the threshold-dependent effects of Pb contamination on soil microbial diversity and denitrifying function. By integrating evidence from studies conducted across contaminated agricultural lands, mining regions, and industrial environments, this review aims to clarify how Pb influences microbial community structure, functional resilience, and nitrogen-transforming processes under varying environmental conditions. Particular attention is given to nonlinear microbial responses, ecological thresholds, and the interplay between soil chemistry and microbial adaptation. Ultimately, understanding these relationships may help inform more ecologically grounded approaches to soil remediation, contamination management, and long-term ecosystem sustainability.