1. Introduction
Eutrophication has become one of the most persistent environmental challenges affecting freshwater ecosystems worldwide. At its most basic level, eutrophication occurs when excessive nutrients—primarily nitrogen and phosphorus—enter aquatic systems, stimulating rapid growth of algae and aquatic plants. Although nutrient enrichment is a natural ecological process, human activities have dramatically accelerated it through agricultural runoff, urbanization, and industrial discharge (Withers et al., 2014; Duncan et al., 2012). The ecological consequences of this imbalance are far-reaching. In many lakes and reservoirs, nutrient enrichment triggers dense algal proliferations that disrupt aquatic food webs, reduce oxygen availability, and degrade water quality (Rabalais et al., 2009; Padedda et al., 2017). Increasingly, such events are not merely ecological concerns but also environmental health hazards.
Among the organisms responsible for these blooms, cyanobacteria have drawn particular attention. These ancient photosynthetic microorganisms thrive in nutrient-rich waters and frequently dominate phytoplankton communities during eutrophic conditions (Sivonen & Jones, 1999). While cyanobacteria play important ecological roles in nitrogen fixation and primary production, many species also synthesize bioactive secondary metabolites known as cyanotoxins. One of the most extensively studied groups of these toxins is microcystins, a class of cyclic peptides that can accumulate in aquatic environments and pose risks to both wildlife and humans (Van Apeldoorn et al., 2007). Monitoring programs worldwide increasingly report the presence of these toxins in bloom-affected waters, underscoring their environmental relevance (Catherine et al., 2017).
Microcystins are structurally diverse compounds, with more than three hundred variants identified to date through modern analytical techniques (Bailiu-Rodriguez et al., 2022). Among them, microcystin-leucine-arginine (MC-LR) is considered the most toxic and widely detected form. These molecules are characterized by a cyclic heptapeptide structure containing a unique ADDA amino acid residue, which contributes to their chemical stability and biological activity (Van Apeldoorn et al., 2007). This structural resilience allows microcystins to persist in aquatic systems despite exposure to ultraviolet radiation, fluctuations in temperature, and microbial degradation processes (Harada et al., 1996). As a result, toxins may remain present in water bodies even after visible cyanobacterial blooms have dissipated.
Environmental persistence is further influenced by interactions between microcystins and sediments. Once released into water bodies, the toxins can bind to particulate matter or accumulate in bottom sediments, creating secondary reservoirs of contamination (Chen et al., 2006). Microbial activity within these sediments can alter toxin concentrations and influence the structure of microbial communities themselves (Ding et al., 2020). Such interactions complicate environmental management because they allow toxins to re-enter the water column long after bloom events, prolonging potential exposure.
The toxicological effects of microcystins are primarily associated with their impact on cellular regulatory systems. These compounds act as potent inhibitors of serine/threonine protein phosphatases, particularly PP1 and PP2A, enzymes essential for maintaining cellular phosphorylation balance (MacKintosh et al., 1990). When these enzymes are inhibited, phosphorylation pathways become dysregulated, leading to cytoskeletal disruption, oxidative stress, and eventually cell death (Dawson, 1998). Cellular uptake of microcystins occurs through organic anion transporting polypeptides, which facilitate toxin entry into hepatocytes and other tissues (Fischer et al., 2005). Experimental studies using animal models have demonstrated that this uptake can result in toxin accumulation within organs and may trigger inflammatory responses and metabolic disturbances (Greer et al., 2018). Histological analyses further reveal that microcystin exposure can damage liver tissues and alter inflammatory pathways (Cao et al., 2019).
Human exposure to microcystins has been documented in several environmental contexts. Individuals may encounter these toxins through ingestion of contaminated drinking water, recreational contact with bloom-affected waters, or inhalation of aerosolized toxins near water bodies (Drobac et al., 2013). Epidemiological investigations have confirmed the presence of microcystins in the blood serum of populations chronically exposed to contaminated water sources (Chen et al., 2009). In extreme cases, acute intoxication events have resulted in severe liver failure and fatalities, as demonstrated by the widely reported hemodialysis incident in Brazil during the 1990s (Jochimsen et al., 1998). Long-term exposure has also been linked to increased risks of liver disease and certain cancers in affected populations (Svircev et al., 2014; Ueno et al., 1996). A growing body of toxicological literature highlights the broader health implications of harmful algal bloom toxins, emphasizing their potential to affect multiple physiological systems (Lad et al., 2022). Laboratory experiments likewise indicate that repeated exposure may induce apoptosis and other pathological responses in animal tissues (Lezcano et al., 2012).
While aquatic exposure pathways have been studied extensively, an emerging concern involves the transfer of microcystins into agricultural environments. Freshwater resources contaminated by cyanobacterial blooms are frequently used for irrigation, particularly in regions experiencing water scarcity. Irrigation with contaminated water can introduce dissolved microcystins directly into agricultural soils and plant systems (Pfister et al., 2011). Groundwater and surface water sources used in agriculture may therefore act as vectors for toxin transport into food production systems (U.S. Environmental Protection Agency, 2021).
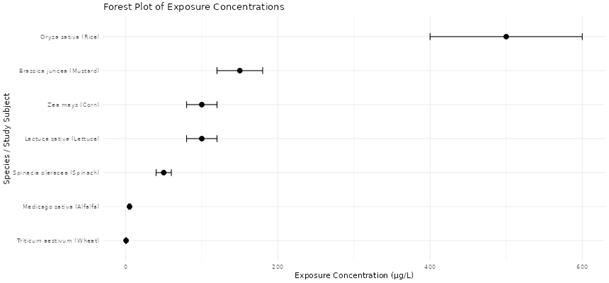
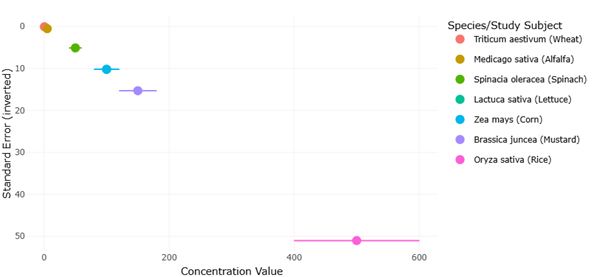
A growing number of studies indicate that agricultural crops are capable of absorbing microcystins from irrigation water. Experimental investigations have demonstrated uptake of toxins in seedlings of several plant species, suggesting that roots can absorb microcystins and translocate them to aerial tissues (Peuthert et al., 2007). Field studies similarly report the presence of microcystins in edible plant tissues when vegetables are irrigated with contaminated water sources (Mohamed & Al Shehri, 2009). The extent of accumulation varies across species and environmental conditions, but leafy vegetables often show higher concentrations due to their physiological characteristics and high transpiration rates.
Beyond accumulation, microcystins may also exert direct toxic effects on plants. Research examining plant physiology has revealed that exposure can interfere with photosynthesis, reduce biomass production, and impair nutrient assimilation (Campos et al., 2021). Early experimental evidence demonstrated that microcystin-LR can inhibit photosynthetic activity in bean leaves, highlighting its potential to disrupt plant metabolic processes (Abe et al., 1996). Similar responses have been observed in rice seedlings exposed to contaminated irrigation water, where growth inhibition and physiological stress responses were reported (Liang & Wang, 2015). Such findings suggest that microcystins may influence not only food safety but also agricultural productivity.
Agricultural practices themselves may also play a role in the distribution of cyanobacterial toxins. Cyanobacteria are sometimes applied as biofertilizers due to their capacity to enhance soil fertility and support sustainable agricultural systems (Singh et al., 2016). Although these microorganisms can improve nutrient cycling, their potential to produce toxins introduces an additional dimension of environmental risk. In bloom-affected regions, water used for crop production may contain cyanobacteria capable of releasing microcystins, thereby increasing the likelihood of contamination within agricultural landscapes (Mutoti et al., 2022).
Despite increasing recognition of these pathways, the environmental behavior and agricultural implications of microcystins remain incompletely understood. Studies examining plant uptake, toxin persistence in soils, and impacts on crop physiology often report inconsistent findings, reflecting differences in experimental design and environmental conditions. Moreover, the interactions between microcystins, soil microorganisms, and plant metabolic pathways add layers of complexity that have yet to be fully resolved.
Given these uncertainties, synthesizing the available scientific evidence is essential for understanding the broader implications of microcystin contamination. Systematic reviews provide an effective framework for integrating diverse research findings and identifying patterns across studies. By critically evaluating published literature, such analyses can clarify exposure pathways, highlight knowledge gaps, and guide future research directions.
The present systematic review therefore examines the occurrence, environmental pathways, phytotoxic effects, and potential human health implications of microcystins within agricultural systems. Particular attention is given to the mechanisms through which these toxins move from aquatic environments into crop production systems and the extent to which they accumulate in edible plants. By integrating findings from environmental science, toxicology, and plant physiology, this review aims to provide a comprehensive perspective on how cyanobacterial toxins intersect with agricultural sustainability and food safety. Ultimately, understanding these dynamics is essential for developing effective monitoring strategies and mitigating risks associated with cyanotoxin contamination in agricultural landscapes.