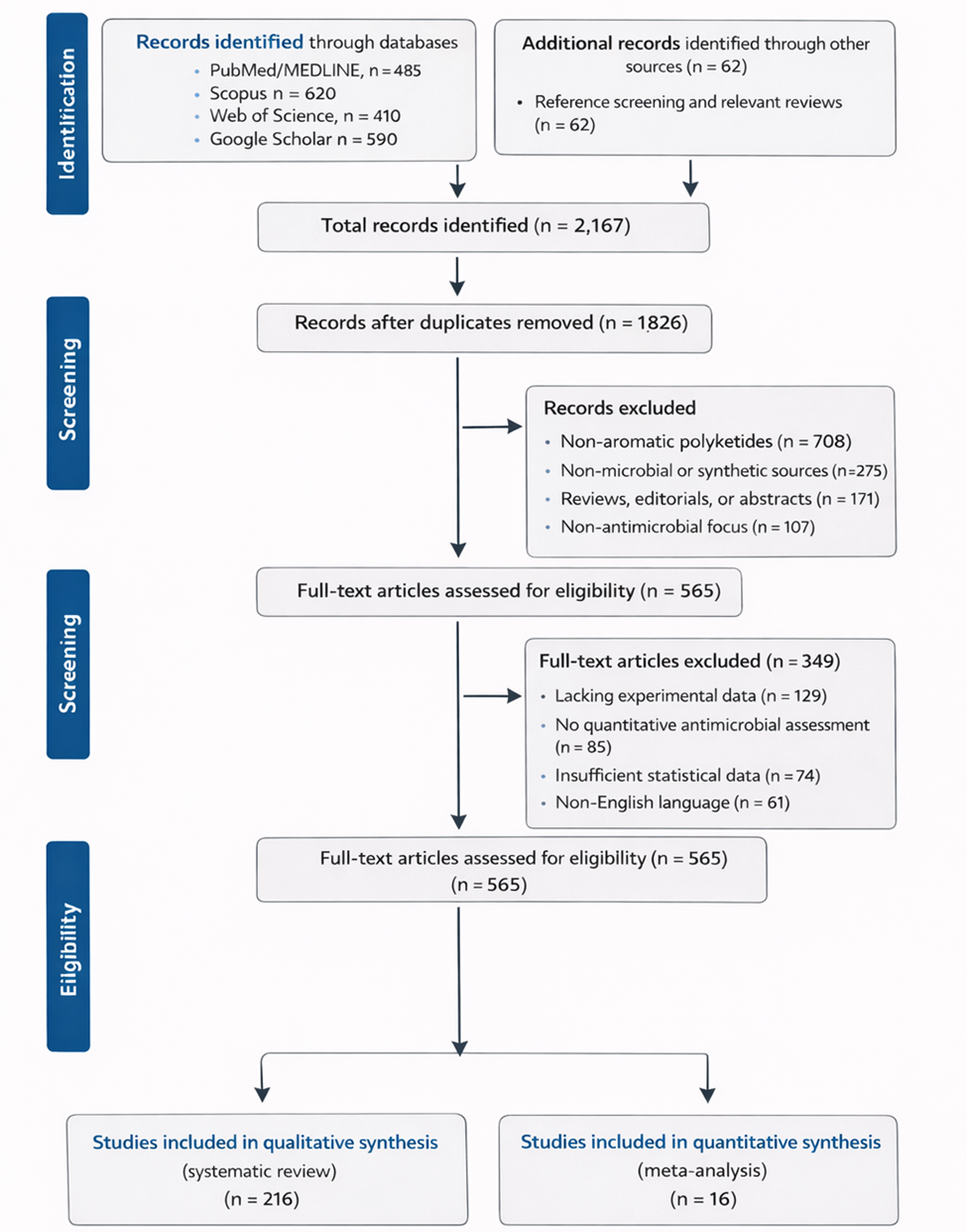
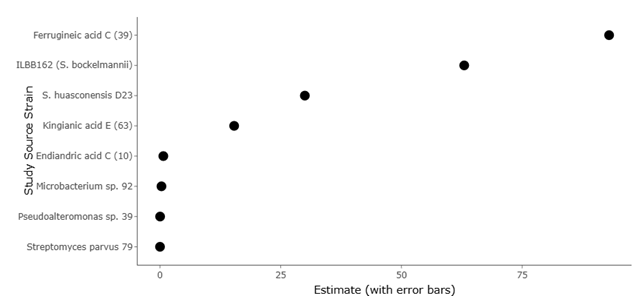
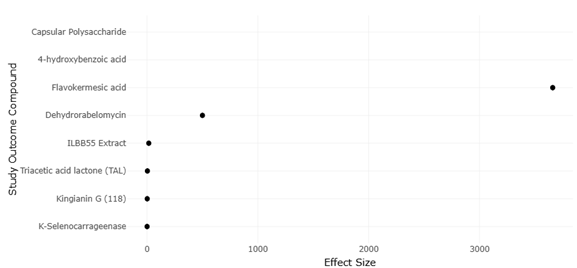
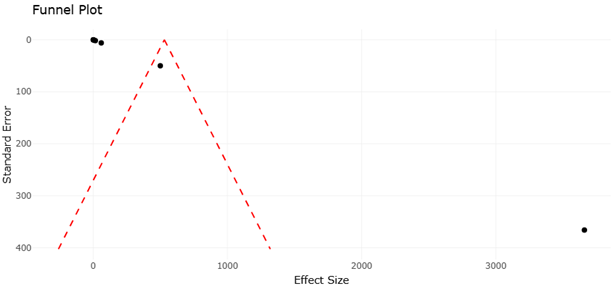
1. Introduction
The rapid rise and worldwide spread of antimicrobial resistance (AMR) represents one of the most pressing challenges confronting modern medicine and global public health. Infections that were once readily cured with standard antibiotics are now increasingly difficult to manage, resulting in prolonged hospital stays, higher treatment costs, and increased mortality. Drug-resistant pathogens compromise routine medical procedures, including surgery, cancer chemotherapy, organ transplantation, and neonatal care, all of which depend heavily on effective antimicrobial prophylaxis and therapy. The drivers of AMR are multifactorial, including misuse and overuse of antibiotics in healthcare and agriculture, as well as the remarkable adaptive capacity of microorganisms (Aslam et al., 2018).
In response to this growing crisis, the search for novel antimicrobial agents with distinct chemical scaffolds and mechanisms of action has become a scientific priority. Historically, natural products have served as the foundation of antibiotic discovery. Microbial secondary metabolites, shaped by millions of years of ecological competition, have yielded structurally diverse and pharmacologically potent compounds. Marine actinomycetes, in particular, have been highlighted as a promising resource for new drug leads due to their unique ecological adaptations and biosynthetic potential (Fenical & Jensen, 2006).
Among microbial producers, actinobacteria—especially members of the genus Streptomyces—have played a dominant role in antibiotic development. Their taxonomy, physiology, and metabolic versatility underpin their extraordinary capacity to synthesize bioactive molecules (Barka et al., 2016). Whole-genome sequencing of environmental Streptomyces isolates has revealed extensive biosynthetic gene clusters encoding diverse secondary metabolites, many of which remain uncharacterized (Alam et al., 2022). Despite this richness, traditional bioactivity-guided screening approaches have frequently resulted in the rediscovery of known compounds, prompting the need for more innovative discovery strategies.
Recent efforts have shifted toward underexplored marine and symbiotic ecosystems, where microorganisms are subjected to distinct selective pressures. Coral reef environments, such as those in the Maldives, harbor complex microbial communities with diverse metabolic capabilities (Palma Esposito et al., 2025). Investigations into sponge-associated bacteria have similarly revealed a wide array of biosynthetic gene clusters encoding potentially novel secondary metabolites (Dat et al., 2023). Marine probiotics, including species of Pseudoalteromonas, have also demonstrated antibacterial and anticancer potential, underscoring the therapeutic promise of ocean-derived microbes (Eze et al., 2023).
At the same time, environmental stressors such as climate change and habitat disruption are reshaping marine ecosystems and influencing microbial diversity (Hodapp et al., 2023). Coral growth anomalies and environmental stress in reef systems have been documented in the Maldives, emphasizing the dynamic and vulnerable nature of these habitats (Bises et al., 2024). Restoration initiatives, including lagoon nurseries and coral gardening strategies, are being implemented to mitigate ecological degradation (Dehnert et al., 2022; Dehnert et al., 2023). Research has also examined the cellular and physiological impacts of microplastic exposure on corals, further highlighting the environmental pressures that shape marine microbial communities (Isa et al., 2024). Innovative biomaterial approaches, such as biodegradable films designed for controlled delivery of protective compounds, have been explored to enhance coral resilience under thermal stress (Contardi et al., 2023). These ecological contexts are relevant because environmental stress can influence microbial secondary metabolism, potentially altering the production of bioactive compounds.
Within the vast repertoire of microbial natural products, polyketides represent one of the most structurally diverse and therapeutically significant classes. Aromatic polyketides, including clinically important anthracyclines, exhibit potent antimicrobial and anticancer activities (Hulst et al., 2022). Their biosynthesis is mediated by polyketide synthases (PKSs), multifunctional enzymatic assemblies that construct complex carbon skeletons through iterative condensation reactions. Type II PKSs, in particular, play a central role in the formation of aromatic frameworks and demonstrate intricate enzymatic coordination (Hertweck et al., 2007). Structural analyses have provided detailed insights into the selectivity mechanisms of highly reducing Type II PKS systems, revealing how subtle conformational features govern product specificity (Du et al., 2020).
Recent structural biology studies have captured high-resolution snapshots of minimal PKS systems, illuminating the molecular basis of octaketide biosynthesis and deepening our understanding of enzymatic assembly lines (Bräuer et al., 2020). Additionally, investigations into uncommon Type II PKSs have uncovered novel catalytic mechanisms responsible for the biosynthesis of aryl polyene pigments, expanding the known diversity of polyketide pathways (Grammbitter et al., 2019). These findings collectively underscore the modular and adaptable nature of PKS enzymes, making them attractive platforms for rational engineering.
Advances in computational biology have further accelerated progress in this field. Machine-learning–based protein structure prediction tools now enable highly accurate modeling of enzyme architecture and protein–protein interactions, facilitating the rational redesign of biosynthetic pathways (Baek et al., 2021). Such technologies complement genome mining approaches by linking genetic information to structural and functional predictions.
Despite the abundance of biosynthetic gene clusters identified through genomic analyses, many remain transcriptionally silent under laboratory conditions. The concept of “cryptic” or “silent” secondary metabolism reflects the vast reservoir of untapped chemical diversity encoded within microbial genomes (Hoskisson & Seipke, 2020). Unlocking these hidden pathways requires innovative strategies, including co-culture techniques, epigenetic modulation, and heterologous expression in optimized host organisms. By activating silent gene clusters, researchers can access previously inaccessible metabolites with potential therapeutic applications.
Metabolic engineering and synthetic biology provide powerful tools to overcome limitations associated with native producers. By reconstructing and optimizing polyketide biosynthetic pathways in tractable hosts, it is possible to enhance yield, modify structural features, and generate novel analogues with improved pharmacological profiles. The integration of genomic data, structural insights, and computational modeling supports a rational approach to pathway refactoring and combinatorial biosynthesis.
The convergence of ecological exploration, genome mining, structural biology, and synthetic biology has revitalized natural product research in the context of antimicrobial resistance. Marine and environmental microorganisms offer a rich and still largely untapped source of biosynthetic diversity. Aromatic polyketides, supported by sophisticated PKS enzymology and modern engineering strategies, stand out as promising scaffolds for next-generation therapeutics. By integrating multidisciplinary evidence and leveraging emerging technologies, the discovery and development of innovative antimicrobial agents can be accelerated to address one of the most formidable health challenges of our time.