3.1 Plastic Pollution as an Environmental and Public Health Crisis: Setting the Stage
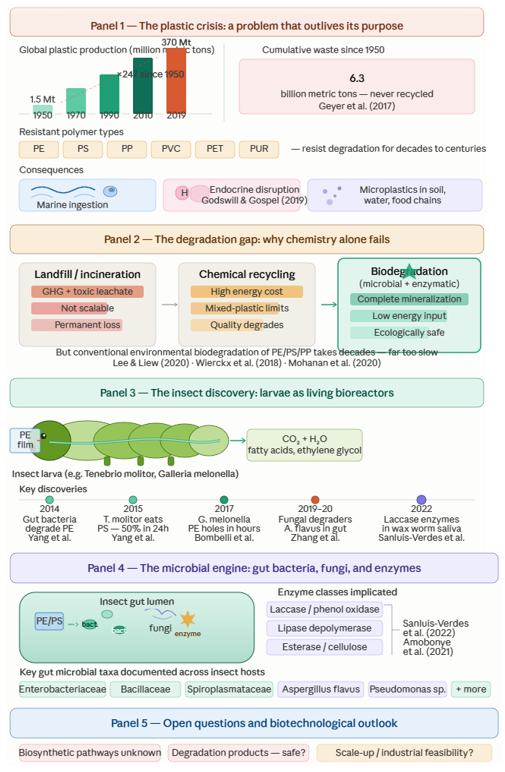
To understand why insect gut microbiomes matter as biodegradation systems, it helps to first appreciate the scale of the problem they are being asked to solve — and that scale is, genuinely, difficult to overstate. Plastic waste has become one of the most pervasive and intractable forms of environmental contamination in the modern era, its reach extending from deep ocean sediments to drinking water supplies to the bloodstream of terrestrial vertebrates. Siddiqui et al. (2024) and Dey et al. (2023) both positioned plastic waste among the defining environmental threats of this century, noting that its accumulation is driven not only by the sheer quantity of material produced but by a structural mismatch between production rates and the global infrastructure available to manage what results. Roughly 80% of plastic waste worldwide ends up in landfills (Landon-Lane, 2018; Paletta et al., 2019) — not because landfilling is considered adequate, but because alternatives are largely absent or underfunded, particularly in the developing economies where production and consumption are growing fastest.
The consequences spread through environmental compartments in ways that compound one another. Landfilled plastics leach into soils, depleting fertility and contaminating groundwater. Improperly managed waste reaches waterways, where it enters the marine environment and persists for timescales measured in centuries — floating, fragmenting, and being ingested by organisms at every trophic level (Kibria et al., 2023; Masud et al., 2019; Paletta et al., 2019). Open burning, deployed when disposal capacity runs out, releases greenhouse gases (GHG), carbon oxides, nitrogen oxides, and a range of additional toxic compounds into the atmosphere, adding an air quality dimension to what began as a solid waste problem (Kibria et al., 2023). Ragossnig and Agamuthu (2021) observed that the physical properties that made plastics commercially attractive — low density, durability, non-degradability — are precisely the properties that make their environmental management so difficult.
3.1.1 The microplastic dimension
The story does not end with macroplastic waste. Over time, and through a range of physical, chemical, and biological processes, larger plastic items fragment into progressively smaller particles — the microplastics that have now been detected in practically every environmental matrix on the planet. Malankowska et al. (2021) drew a useful conceptual distinction here: primary microplastics are manufactured intentionally at small size for use in personal care products, industrial abrasives, and medical applications, while secondary microplastics arise from the environmental breakdown of larger plastic items through exposure to UV radiation, thermal oxidation, mechanical shear, wave action, and biofilm development (Andrady, 2011). Both pathways lead to the same outcome — persistent, mobile, sub-millimetre particles that do not settle readily, are not captured by most water treatment processes, and accumulate in living tissues.
The geographical reach of microplastic contamination is, at this point, essentially global. Desforges et al. (2015) documented microplastic ingestion by zooplankton in the northeast Pacific. Gasperi et al. (2018) reported microplastics in urban air samples, raising the prospect of inhalation as an exposure route. Kosuth et al. (2018) found anthropogenic particle contamination in tap water, beer, and sea salt across multiple countries. Schymanski et al. (2018) detected microplastic release from packaging into bottled mineral water. More recently, studies across India, the United States, Poland, Germany, the United Kingdom, Australia, Sweden, and Denmark confirmed microplastic presence in groundwater and drinking water supplies (Khant & Kim, 2022). The contamination of freshwater ecosystems — rivers, lakes, ponds, canals, and estuaries — via domestic wastewater discharge and the degradation of single-use plastic items has been documented across Asia, Europe, and the Americas (Sarijan et al., 2021; Goveas et al., 2023). The classification of microplastics by chemical composition — PS, PE, PP, PVC, PET, and polyamide (PA), among others — identified across these matrices aligns closely with the dominant polymer types that insect larvae have been shown to degrade (Siddiqui et al., 2024).
What makes microplastic contamination particularly alarming, biologically, is not simply its ubiquity but its demonstrated capacity to enter food chains at the lowest trophic levels and accumulate upward. Plastic particles are also vectors for adsorbed chemical contaminants — persistent organic pollutants, heavy metals, and plastic-specific additives including phthalates and bisphenols — which can leach into tissues at points of ingestion or during digestive processing. Sharma and Chatterjee (2017) and Ng et al. (2018) documented these chemical transfer dynamics across agroecosystems and marine environments respectively. The implications for human health via dietary exposure to contaminated seafood and water remain an area of active investigation, though the evidence for endocrine disruption via plastic-associated compounds is already well established (Godswill & Gospel, 2019).
3.2 Why Conventional Degradation Approaches Are Insufficient
Understanding the case for insect gut microbiome-based biodegradation requires understanding what the alternatives offer — and, just as importantly, what they do not. The degradation of synthetic plastics in the environment can, in principle, proceed through several mechanisms: abiotic pathways including photochemical, thermal, and mechanical degradation; conventional biological degradation by free-living environmental microorganisms; and managed engineering approaches including chemical recycling, mechanical recycling, and incineration. Each has genuine utility in specific contexts. None, individually or collectively, is remotely adequate to address the current scale of plastic waste accumulation.
Gewert et al. (2015) described the abiotic degradation pathways operating on floating marine plastics — UV-mediated chain scission, oxidative thermal degradation, and physical fragmentation — concluding that while these processes initiate surface oxidation and molecular weight reduction over timescales of decades, they do not achieve mineralization. What they produce, in most cases, is more microplastics. Thermal degradation in ambient terrestrial environments is similarly slow; photodegradation is somewhat more efficient outdoors but still operates over years to centuries for bulk polymer, and is negligible in buried or aquatic contexts where UV penetration is limited (Gewert et al., 2015). Akbay and Ozdemir (2016) confirmed that natural abiotic degradation of conventional plastics is, in practical terms, too slow to prevent continued accumulation under current production rates.
Plastic waste management approaches — mechanical recycling, chemical recycling, and incineration — each address portions of the problem but carry significant constraints. Mechanical recycling, which reprocesses thermoplastic waste into new products, suffers from cumulative quality degradation across processing cycles, mixed-material incompatibility, and collection and sorting infrastructure limitations. Chemical recycling can, in principle, recover 61 different plastic types into monomers and other useful chemical feedstocks (Rahimi & Garcia, 2017), but remains economically marginal for most polymer types at current scale, and energy-intensive across all. The current global treatment breakdown — approximately 77% reclamation (landfill or open disposal), 13% incineration, and 10% mechanical and chemical recycling (Briassoulis, 2006) — reflects not what is desirable but what is actually happening, and it is far removed from a sustainable equilibrium. Incineration, meanwhile, destroys material value while generating GHG and, depending on incineration conditions and plastic composition, potentially toxic combustion products including carcinogens such as ketones and related compounds (Briassoulis, 2006).
Biological degradation — microbial, enzymatic, or organism-mediated — is the approach that, in theory, most closely approximates what natural carbon cycling achieves for organic materials. The challenge, as discussed in the introduction and elaborated below, is that conventional environmental microbial degradation of polyolefin plastics (PE, PS, PP) proceeds at rates that are, in most terrestrial and aquatic settings, measured in decades to centuries (Mohanan et al., 2020; Lee & Liew, 2020). The enzymes capable of efficiently attacking the carbon–carbon backbone bonds of polyolefins are simply not abundant in most environmental microbial communities, because these bonds — unlike the ester linkages of PET or the ether linkages of some polyurethanes — have no close analogue in natural biochemistry. It is precisely this biochemical gap that makes the discovery of efficient plastic-degrading enzymatic systems in insect gut environments so significant, and so scientifically interesting.
3.3 Insects as Biological Hosts for Plastic Degradation: A Cross-Species Overview
The observation that certain insect larvae consume, and in some cases substantially degrade, synthetic plastic polymers has now been documented across at least ten species drawn from two insect orders — Lepidoptera and Coleoptera (Table 1). The breadth of this distribution is itself informative: it suggests that plastic degradation capacity is not an isolated evolutionary novelty confined to
Table 1. Documented insect species with confirmed plastic biodegradation capacity, associated gut symbiotic microorganisms, plastic substrate types, quantitative degradation efficiencies, and key mechanistic evidence (1997–2024).
|
Insect species
|
Types of plastics
|
Symbiont microbes
|
References
|
|
Galleria melonella
|
PE, PS
LDPE
LDPE
PS
PS
|
Intestinal bacteria-Bacillus and Serratia.
Gut Microbiome-Acinetobacter, closcibacterium, Corynebacterium Curvibacter, Enhydrobacter and Staphylococcus genera
Gut microbiome
Gut microbiota
Intestinal bacteria, Massilia sp. FS 1903
|
Lou et al., 2020
Latur et al., 2021
Rijasee et al., 2021
Wang et al., 2022
Jiang et al., 2020
|
|
Achroia grisella
|
HDPE
|
-----------
|
Kundungal et al., 2019
|
|
Plodia interpunctella
|
PE
PE
|
Enterobacter asburiae YT1 and Bacillus sp. YP1
Meyerozyma guilliermondii ZJC1
(MgZJC1) and Serratia marcescens
ZJC2 (SmZJC2)
|
Yang et al. 2014
Lou et al., 2022
|
|
Tenebrio molitor
|
PE, PS
PS
Polyether-Pu foam
PE
PS
PS
PVC
|
Gut microbiome-Citrobacter sp. and Kosakonia sp.
Gut microbiome-eight unique bacterial species
Gut Microbiome-the families
Gut microbione
Gut microbiom-Enterococcus, Enterobacteriaceae- Escherichia-Shigell, and Lactoco-ccus.
Cronobacter Sukazakii and Lactococcus garvieae
Gut microbiome
|
Brandon et al., 2018
Brandon et al., 2021
Liu et al., 2022
Bulak et al., 2021
Jiang et al., 2021a
Bae et al., 2021
Peng et al., 2020a
|
|
Tenelerio obscurs
|
PS
|
Intestinal bacteria-Enterobacteriaceae,
Spiroplasmatceae, and Enterococcaceae
|
Peng et al., 2019
|
|
Tribolium confusum
|
PS, PE and EVA (ethyl vinylacetate
|
------
|
Abdulhay 2020
|
|
Tribolium castaneum
|
PS
|
Intestinal bacterium, Acinetobacter
|
Wang et al., 2020
|
|
Uloma spp.
|
PS
|
Gut micro biota
|
Kundungal et al., 2021.
|
|
Zophobas atratus
|
PS
PS
PS foam
PS, LDPE
LDPE, EPS
|
Gut microbiome-Pseudomonas sp., EDB1
Bacillus sp. EDA4 and Brevibacterium sp. EDX
Gut microbiome-Enterococcus.
Gut microbiome
Gut microbiota and microbial functional enzymes
Gut microbiota
|
Arunrattiyakorn et al., 2022
Jiang et al. 2021b
Yang et al., 2020
Peng et al., 2022
Peng et al., 2020b
|
Table 2. Additional insect and arthropod species reported to penetrate, consume, or interact physically with synthetic plastic packaging materials, for which systematic gut microbiome characterisation or controlled biodegradation data remain limited or absent (1955–2019).
|
Insect species
|
Plastic types
|
References
|
|
Ephestia cautella
Lasioderma serricorne
Oryzaephilus surinamensis
Callosobruchus maculatus
Rhizopertha dominiea
Sitophilus oryza
Sphaeroma terebrans
Stegobium Paniceum
|
PVC, PP
PP, PE, PEST
PE
PE
PP, PE, PEST
PP, PE, PEST
PS
PS
|
Graham Bowditch, 1997
Riudavets et al., 2007
Al 1993
Al, 1993
Graham Bowditch 1997
Riudavents et al., 2007
Davidson, 2012
Davidson, 2012
|
Table 3. Structured literature search summary across six scientific databases for the systematic narrative review on insect gut microbiome-mediated plastic biodegradation, covering publications from 1955 to 2024.
|
Database
|
Search period
|
Search terms applied
|
Approximate records retrieved
|
|
PubMed/MEDLINE
|
1993–2024
|
Cluster A + B + C
|
~180
|
|
Web of Science
|
1993–2024
|
Cluster A + B + C
|
~240
|
|
Scopus
|
1993–2024
|
Cluster A + B + C
|
~210
|
|
ScienceDirect
|
1993–2024
|
Cluster A + B + C
|
~90
|
|
Wiley Online Library
|
1993–2024
|
Cluster A + B + C
|
~60
|
|
SpringerLink
|
1993–2024
|
Cluster A + B + C
|
~75
|
|
Total after deduplication
|
|
|
~65 included
|
a single lineage, but a functional property that has emerged, or been co-opted, across phylogenetically diverse gut microbiome communities residing in taxonomically disparate hosts. Bilal et al. (2021) listed seven plastic types demonstrably degraded by insects — PE, PS, PVC, PP, polyphenylene sulfide (PPS), ethylene vinyl acetate (EVA), and extruded polystyrene (EPS) — and the range has continued to expand. A comprehensive summary of documented insect species, plastic types, and associated symbiotic microorganisms is provided in (Table 1), with additional insect species whose degradation capacity remains insufficiently characterized listed separately in (Table 2).
What is worth emphasizing before moving to individual species accounts is the interpretive challenge that this literature presents. Degradation efficiencies reported across different studies are rarely directly comparable: they differ in how degradation is measured (mass loss, molecular weight reduction, isotopic tracing, CO₂ evolution), in experimental duration, larval number, and temperature, and in whether the larvae were maintained on a pure plastic diet or a mixed diet supplemented with conventional food. Chen et al. (2020) argued that the synergistic combination of insect enzymatic activity and gut microorganisms enables plastic polymer breakdown in ways that neither component achieves alone, and this mechanistic complexity makes inter-study comparison difficult. The species-by-species synthesis below should be read with this caveat in mind.
3.3.1 Galleria melonella (greater wax moth)
Among all the insect species documented in this field, Galleria melonella occupies something like a founding position — it was the subject of the finding that first generated widespread scientific and public interest in insect-mediated plastic degradation, and it has remained one of the most intensively studied systems since. The larval biology of G. melonella provides a plausible biochemical rationale for its PE-degrading capacity: as a parasite of bee colonies, the larvae feed naturally on beeswax, a complex mixture of long-chain saturated hydrocarbons whose structural chemistry is broadly analogous to that of polyethylene (Bombelli et al., 2017; Khyade, 2018; Grove & Villanueva, 2021). The hypothesis — still not definitively proven, though increasingly well supported — is that the biochemical machinery evolved for wax metabolism has been recruited, or at least partially adapted, for the oxidative breakdown of PE polymer chains.
Bombelli et al. (2017) reported the landmark observation that G. melonella larvae could produce visible pitting in commercial PE films within an hour of contact, and that approximately 100 larvae caused a loss of 92 mg of PE over 12 hours under ambient conditions — a rate far exceeding anything previously reported for a biological system (Groh, 2017; Weber et al., 2017). The mechanism, initially attributed to gut microbial activity, was subsequently shown to be more complex and partly host-driven. Peydaei et al. (2020) demonstrated through salivary gland proteomics that PE exposure induces changes in the larval salivary proteome, with accumulation of carbonyl groups at pitted surface areas indicating oxidative modification of the polymer at points of larval contact. Le Moine et al. (2020) showed that lipid oxidative enzyme activities are significantly upregulated in larvae maintained on PE as a dietary substrate, and that the larvae can redirect their lipid homeostasis machinery to extract metabolic energy from the polymer. The identification by Sanluis-Verdes et al. (2022) of two phenol oxidase enzymes — classified within the laccase family and designated PEases — in wax worm saliva brought the host-derived enzymatic contribution into sharp focus: these enzymes could initiate oxidative cleavage of PE polymer chains at room temperature within hours, independently of gut microbial activity.
The gut microbial contribution to G. melonella PE degradation is equally well documented, and operates in parallel to rather than instead of the salivary pathway. Ren et al. (2019) isolated Enterobacter sp. D1 from the gut of G. melonella larvae and demonstrated a 6.1–10.7% reduction in PE mass over 28 days in the presence of this bacterium combined with the gut-associated fungus Aspergillus flavus. Zhang et al. (2020) characterized A. flavus as an independent PE-degrading organism capable of reducing high-density polyethylene (HDPE) molecular weight in 28-day incubations through extracellular enzymatic activity. Lou et al. (2022) isolated Bacillus sp. and Serratia sp. from G. melonella gut contents and reported mass losses of 0.88 g (PE) and 1.95 g (PS) respectively over 21 days in isolated culture — rates that, while modest in absolute terms, are substantial relative to what free-living environmental bacteria achieve under comparable conditions.
The broader gut microbiome picture for this species, as captured in (Table 1), includes Acinetobacter, Clostridium, Corynebacterium, Curvibacter, Enhydrobacter, and Staphylococcus genera, documented by Latour et al. (2021), as well as the Massilia sp. FS1903 strain reported by Jiang et al. (2021b). The compositional diversity of this community is striking, and it raises a question that remains only partially resolved: which taxa are primarily responsible for enzymatic polymer attack, and which are secondary colonizers benefiting from the breakdown products generated by others? Lou et al. (2020) found that when G. melonella larvae were fed PE, the intestinal microbiome increased in both species richness and evenness relative to controls on a conventional diet — consistent with the idea that plastic-derived oligomers and fatty acid intermediates provide novel carbon sources supporting a broader microbial community. Wang et al. (2022) contributed an important mechanistic nuance: their demonstration that G. melonella larvae could achieve complete digestion and biodegradation of PS microplastics in vivo, with evidence for this capacity persisting even when gut microbiota were substantially suppressed, echoes the Kong et al. (2019) finding that some aspects of G. melonella's plastic-metabolizing capability may reside in the host hologenome rather than exclusively in the gut microbiome. The relationship between host-intrinsic and microbiome-mediated degradation in this species remains genuinely unresolved.
3.3.2 Achroia grisella (lesser wax moth)
Achroia grisella, a close relative of G. melonella in the family Pyralidae, has attracted considerably less experimental attention, but the available evidence suggests a meaningful PE-degrading capacity, again plausibly linked to wax-metabolizing biochemistry. Kundungal et al. (2019) documented HDPE degradation by A. grisella larvae, using the appearance of new carbonyl and alcoholic functional groups in larval frass — detectable by FTIR spectroscopy — and increases in unsaturated hydrocarbon content as chemical indicators of oxidative polymer modification. While no symbiotic microorganisms were characterized in that study (Table 1), the biochemical signature of degradation is consistent with the oxidative mechanisms documented in G. melonella. Chalup et al. (2018) published the first report of A. grisella larvae consuming agricultural silo bags — multilayer PE-based structures including an anti-UV protective layer — in a real-world agricultural setting in Argentina, confirming that the laboratory observations have ecological correlates outside controlled conditions.
3.3.3 Plodia interpunctella (Indian meal moth)
The Indian meal moth occupies an interesting position in this literature as one of the species for which the gut bacterial contribution to PE degradation was most clearly established in early work. Bowditch (1997) first documented P. interpunctella larvae penetrating polyethylene packaging films and causing physical damage — an observation that predates the mechanistic period but established the basic fact of larval–plastic interaction. Yang et al. (2014) subsequently isolated Enterobacter asburiae YT1 and Bacillus sp. YP1 from the gut of P. interpunctella larvae and demonstrated independent PE-degrading activity for both strains in culture, providing the first direct microbial evidence for gut bacteria as degradation agents in a pyralid moth species.
Later work extended the microbiome characterization considerably. Lou et al. (2022) isolated Meyerozyma guilliermondii ZJC1 (MgZJC1) and Serratia marcescens ZJC2 (SmZJC2) from P. interpunctella larvae and demonstrated that the insect combined with these two symbiont microorganisms caused a 15.87% loss in PE mass over 60 days — a rate that, while lower than the fastest reports for T. molitor or G. melonella, is consistent with meaningful biological degradation activity over practical timescales. The isolation of a yeast (M. guilliermondii) alongside a bacterium (S. marcescens) as co-degraders from the same gut environment is particularly interesting, as it points to a multi-kingdom microbial consortium operating within the gut lumen — a complexity that simpler single-isolate studies tend to obscure.
3.3.4 Corcyra cephalonica (rice meal worm)
Corcyra cephalonica is, in some respects, one of the more methodologically informative cases in this literature, because the available studies directly tested the contribution of gut microbiota to LDPE degradation using an antibiotic manipulation design. Kesti and Thimmappa (2019) and Kesti and Sharana (2019) found that C. cephalonica larvae caused a 21% loss in LDPE mass when fed on LDPE without antibiotic treatment, and a 25% loss when antibiotics were co-administered — a counterintuitive result that the authors interpreted as potentially reflecting changes in larval behaviour, feeding rate, or host enzymatic activity under microbiome suppression. The finding is difficult to reconcile straightforwardly with the dominant model of microbiome-dependent degradation, and illustrates the interpretive complexity that antibiotic manipulation experiments can generate when host and microbial contributions are not cleanly separable.
3.3.5 Tenebrio molitor (yellow mealworm)
Tenebrio molitor is, by a considerable margin, the most extensively studied insect in the context of plastic biodegradation, and the body of evidence for this species is correspondingly rich — and mechanistically detailed in ways that the literature on some other species is not. The range of plastic types degraded by T. molitor larvae is broader than for any other species: documented substrates include PS (Yang et al., 2015a, 2015b; Yang et al., 2018; Jiang et al., 2021a), PE including LDPE, LLDPE, and HDPE (Brandon et al., 2018; Yang et al., 2022), PP (Yang et al., 2021), PVC (Peng et al., 2020a), and polyether-polyurethane foam with a reported degradation efficiency of 67% — one of the highest values in the literature for any insect–plastic combination (Liu et al., 2022).
The PS system in T. molitor has been particularly well characterized mechanistically. Yang et al. (2015a) used isotopic tracing (¹³C-labelled PS) to demonstrate complete mineralization of ingested PS to CO₂ — not merely mass loss, but genuine depolymerization to inorganic end-products — with approximately 47.7% of ingested PS carbon recovered as ¹³CO₂. That same isotopic tracing approach confirmed that the remainder was incorporated into larval biomass, indicating metabolic assimilation rather than excretion. Yang et al. (2015b) followed with a companion paper specifically establishing the gut microbiome's role in this process: larvae treated with gentamicin, which suppresses intestinal bacterial activity, lost the ability to degrade PS — a finding that positioned the gut microbiome as essential, not merely contributory. Brandon et al. (2018) documented that approximately 49% of ingested PE was metabolized to CO₂ in T. molitor larvae fed PE as a sole carbon source, and characterized the gut microbiome shifts accompanying this plastic feeding — notably the enrichment of Citrobacter sp. and Kosakonia sp. as dominant genera in PE-fed larvae relative to controls. Brandon et al. (2021) extended this analysis to show that the T. molitor gut microbiome produces emulsification factors that enhance the bioavailability of polystyrene to microbial degraders, reframing the degradation process as involving active modification of substrate physical properties by microbial community secretions.
The enzyme profile of T. molitor gut contents under plastic feeding has been investigated by Przemieniecki et al. (2020), who documented induction of esterase and cellulose-associated hydrolase activities in larvae fed PE and PS relative to cellulose controls — providing direct enzymatic evidence for microbiome-driven metabolic adaptation to synthetic polymer substrates. Yang et al. (2021) confirmed microbiome-dependent depolymerization of PP by both T. molitor and Zophobas atratus via a variety of gut microbial taxa, reinforcing the picture of a flexible, compositionally dynamic gut community capable of adapting its degradative repertoire to different polymer chemistries.
An important practical dimension of T. molitor biology in this context is the well-documented effect of diet supplementation. Brandon et al. (2018) and Kundungal et al. (2019) both reported that larvae maintained on mixtures of plastic and conventional food (typically wheat bran) showed higher degradation rates than those on plastic alone — a finding that Yang et al. (2018, 2023) elaborated in terms of larval growth, survival, and microbiome diversity. The implication is that optimizing the plastic-to-nutrient ratio in larval diets, rather than imposing plastic as the exclusive energy source, may be essential for sustaining both degradation activity and larval health over extended exposure periods — a consideration that has obvious relevance to any future scale-up application.
3.3.6 Zophobas atratus (superworm)
Zophobas atratus larvae have demonstrated degradation capacity across an impressive range of plastic types, including PE, PS (Kim et al., 2020), PPS foams (Li et al., 2020), EPS, and LDPE (Peng et al., 2020b, 2022). The gut microbiome characterization for this species, summarized in (Table 1), includes Pseudomonas sp. EDB1, Bacillus sp. EDA4, and Brevibacterium sp. EDX, isolated by Arunrattiyakorn et al. (2022) from gut contents of PS-fed superworms and demonstrated to degrade PS in isolated culture. Kim et al. (2020) documented PS degradation by Pseudomonas sp. isolated from the Z. atratus gut, while Yang et al. (2020) confirmed that PS biodegradation and mineralization by superworms is gut microbiome-dependent. Peng et al. (2020b, 2022) made a notable observation regarding PE and PS degradation in this species: while both polymers showed measurable degradation, PS breakdown products were not recoverable in larval frass, suggesting more complete metabolic assimilation of PS degradation intermediates compared to PE — a difference that may reflect the aromatic character of PS and the different enzymatic pathways required for its initial depolymerization.
3.3.7 Tribolium castaneum (red flour beetle)
The red flour beetle has been documented degrading polystyrene by Wang et al. (2020), who isolated Acinetobacter sp. from larval gut contents and reported a 12.4% loss in PS mass weight with corresponding molecular weight reduction over 60 days. Fabreag and Familara (2019) provided additional documentation of T. castaneum larvae consuming and biotically processing extruded polystyrene foam (EPS) blocks. The Acinetobacter genus has not been associated with plastic degradation across many other insect species, making this finding interesting from the perspective of understanding the diversity of functional microbial lineages capable of polymer-degrading activity. The contribution of Acinetobacter to PS depolymerization in this system warrants deeper enzymatic characterization.
3.3.8 Tribolium confusum (confused flour beetle)
Abdulhay (2020) reported that T. confusum larvae caused mass losses of 46.84%, 51.92%, and 2.9% in PE, PS, and EVA respectively over 30 days — making this species one of the more effective degraders per unit time in the available literature. The observation that larval mass increased over the experimental period, despite plastic being the primary dietary substrate, led Abdulhay (2020) to suggest that plastic materials serve not only as energy sources but may fulfil additional metabolic or structural roles in larval physiology — an interpretation that remains speculative and would benefit from controlled metabolic studies.
3.3.9 Uloma spp.
Kundungal et al. (2021) reported the remarkable finding that Uloma sp. larvae could not only consume PS but complete their entire life cycle — through egg, larval, pupal, and adult stages — on a polystyrene-only diet, with degradation improving in the second generation of PS-fed insects. Degradation efficiency reached 37.14 mg PS per 100 larvae per day. The suggestion that gut microbiota were primarily responsible for PS biodegradation in this species is consistent with the general pattern, though no specific taxa were isolated or characterized in that study (Table 1). The multi-generational component of this finding is intriguing: it raises the possibility that prolonged exposure to plastic as a dietary substrate selects for gut microbiome communities with enhanced degradation activity — a form of host-associated microbiome adaptation that could be experimentally exploited.
3.3.10 Tenebrio obscurus (dark mealworm)
Tenebrio obscurus, a congeneric species to the much-studied T. molitor, has been documented by Peng et al. (2019) consuming PS at an average rate of 32.44 mg per 100 larvae per day — comparable to some T. molitor values and demonstrating that plastic degradation capacity is not exclusive to the yellow mealworm within the Tenebrio genus. The gut bacterial community responsible was attributed to members of the families Enterobacteriaceae, Spiroplasmataceae, and Enterococcaceae (Peng et al., 2019) — a compositional signature that overlaps partially but not entirely with that of T. molitor, suggesting that while taxonomically related hosts share some gut community architecture, they are not interchangeable in terms of microbial composition or, presumably, degradation mechanism.
Beyond these ten primary species, a further group of stored-product insects and wood-boring crustaceans has been documented interacting with plastic materials — principally by penetrating plastic packaging — though controlled degradation data are more limited (Table 2). These include Ephestia cautella, Lasioderma serricorne, Oryzaephilus surinamensis, Callosobruchus maculatus, Rhizopertha dominica, Sitophilus oryzae, Sphaeroma terebrans, and Stegobium paniceum (Bowditch, 1997; Riudavets et al., 2007; Gerhardt & Lindgren, 1955; Cline, 1970; Davidson, 2012). Whether these species harbour gut microbiota with comparable enzymatic capabilities to those described above remains to be investigated, and represents a meaningful gap in the current understanding of plastic-degrading biodiversity within the insect kingdom.
3.4 The Biochemical Mechanism: A Synergistic Six-Step System
Having surveyed the species-level evidence, it is worth stepping back to consider the degradation mechanism as a unified system — because the species-by-species perspective, while useful for capturing diversity, can obscure the shared biochemical logic underlying plastic breakdown across all of these hosts. Yang et al. (2022) synthesized the available mechanistic evidence into a six-step framework that, while likely simplified relative to the actual complexity of gut biochemistry, provides a useful organizing structure (Figure 1).
The sequence begins — necessarily — with physical fragmentation. Step 1 involves mechanical comminution of plastic material by insect mouthparts and mandibles, reducing bulk polymer to particles small enough to transit the gut lumen and present accessible surface area to microbial and enzymatic attack. This mechanical size reduction is not trivial: the surface-to-volume ratio of plastic fragments determines the rate of enzymatic access to polymer chains, and insects with powerful chewing mouthparts can generate particles far smaller than those produced by most environmental weathering processes on comparable timescales.
In Step 2, gut microbes adhere to plastic particle surfaces and initiate surface erosion — a process that has been visualized directly by scanning electron microscopy in several studies and leaves characteristic surface morphology changes including pitting, cracking, and loss of surface hydrophobicity that can be detected by water contact angle measurements and FTIR spectroscopy. Steps 3 and 4 represent the enzymatic core of the process. In Step 3, depolymerization of plastic into oligomeric fragments begins through oxidative or hydrolytic enzymatic attack, with the enzyme complement contributed by both the insect host and its gut microbiome. In Step 4, the insect host secretes or otherwise contributes bio-emulsifying substances — possibly surfactant-like molecules analogous to those used in lipid digestion — that increase the physical accessibility of polymer surfaces to microbial enzyme attack, essentially acting as a wetting agent for what is otherwise a hydrophobic substrate. Brandon et al. (2021) provided direct evidence for this emulsification function in T. molitor, demonstrating that gut microbiome enrichments from PS-fed larvae showed enhanced respiration on PS in the presence of emulsification factors secreted by the host gut. Steps 5 and 6 involve the progressive breakdown of oligomers through bond cleavage into short-chain fatty acids, which are then assimilated through standard insect metabolic pathways — explaining how plastic-fed larvae can sustain growth and complete development on polymer-only diets, at least for the species where this has been documented.
The antibiotic inhibition experiments conducted across multiple species — notably the gentamicin treatments in T. molitor (Yang et al., 2015b) and similar designs in other hosts — provide the most direct evidence that gut bacteria are functionally essential to this sequence, not merely incidentally present. Suppression of gut bacterial activity consistently reduces or eliminates measurable plastic degradation, positioning the microbiome as a necessary rather than sufficient component of the overall system. At the same time, the demonstration of host-derived enzymatic contributions in G. melonella — particularly the salivary PEases characterized by Sanluis-Verdes et al. (2022) — complicates any simple narrative that reduces this to a purely microbial story. It seems more accurate to say that insect gut environments create a unique set of physical and chemical conditions — mechanical comminution, controlled pH and temperature, emulsification factors, enzymatic substrate pre-oxidation — that collectively enable a resident microbial community to achieve degradation rates impossible in any open environmental context.
3.5 Gut Microbial Taxa and Enzyme Classes: The Biochemical Agents
The microbial taxa documented as plastic-degrading agents in insect gut environments span a taxonomically and functionally diverse range, summarized across species in (Table 1). Rather than cataloguing them exhaustively — which the table already does — it is more useful here to draw out the cross-cutting patterns.
Within the bacteria, three broad functional categories of isolate emerge from the literature. The first comprises Gram-negative enteric bacteria of the family Enterobacteriaceae, represented principally by Enterobacter sp. (Yang et al., 2014; Ren et al., 2019), Citrobacter sp. (Brandon et al., 2018), Kosakonia sp. (Brandon et al., 2018), Serratia marcescens (Lou et al., 2022), and Cronobacter sakazakii (Bae et al., 2021). These are gram-negative, facultatively anaerobic organisms well suited to the variable oxygen tension of the insect gut lumen, and several genera within this family have established reputations as producers of extracellular depolymerases and lipases in other biotechnological contexts. The second category includes Gram-positive spore-forming bacteria of the family Bacillaceae, represented by Bacillus sp. YP1 (Yang et al., 2014; Lou et al., 2022) — organisms known for their broad extracellular enzyme repertoire and tolerance of challenging physical conditions. The third comprises Pseudomonas sp. (Kim et al., 2020; Li et al., 2020), Gram-negative gammaproteobacteria with well-documented capacity for degrading aromatic and aliphatic hydrocarbons in other environmental contexts, and whose presence in insect gut communities degrading both PE and PS is consistent with that biochemical background.
Within the fungi, Aspergillus flavus is the most thoroughly characterized plastic-degrading gut fungus identified to date. Zhang et al. (2020) demonstrated its capacity for HDPE molecular weight reduction via extracellular enzymatic activity over 28 days. The presence of a yeast (Meyerozyma guilliermondii) as a co-degrader in P. interpunctella gut contents (Lou et al., 2022) adds another fungal lineage to the roster, though its enzymatic mechanism has not been characterized.
Turning from taxa to mechanisms, Amobonye et al. (2021) provided the most comprehensive synthesis of the enzyme classes implicated in microbial plastic degradation, identifying lipase depolymerases, proteinases, esterases, urease, chitinase, and dehydrase as among the extracellular enzymes produced by plastic-degrading bacteria — all enzymes that initiate depolymerization by attacking surface-accessible chain or ring structures and reducing bulk polymers to shorter oligomers that can be transported into cells for further oxidation. The specific enzyme most thoroughly characterized in the insect gut context remains the phenol oxidase/laccase enzymes identified by Sanluis-Verdes et al. (2022) in G. melonella saliva, which occupy a mechanistically important position as initiators of oxidative chain cleavage in PE. Przemieniecki et al. (2020) documented esterase and cellulose-related hydrolase induction in T. molitor gut contents under PE and PS feeding. For PET specifically — a polymer with ester linkages more amenable to enzymatic attack than the carbon–carbon bonds of polyolefins — PET hydrolase activity is required to initiate chain modification, and Kawai et al. (2019) and Taniguchi et al. (2019) reviewed the state of PETase characterization in detail, though the insect gut context for PET degradation is less well documented than for PE and PS.
The honest assessment of the enzyme literature is that it remains fragmentary. Many degradation studies report activity outcomes (mass loss, molecular weight reduction) without identifying the responsible enzymes; many enzyme studies characterize activity in isolated bacterial cultures without establishing whether the same activity operates in the intact gut environment. Connecting these two levels of evidence — from organism-level degradation phenotype to specific enzyme mechanism — is the most pressing unmet need in this field, and the area where metagenomics and proteomics approaches have the greatest potential to accelerate understanding.
3.6 Cross-Species Comparisons, Confounding Variables, and Interpretive Cautions
Any attempt to compare degradation efficiencies across species and studies runs into the methodological heterogeneity problem described briefly in Section 3.3. It is worth being explicit about what this means in practice. A reported mass loss of 50% in T. molitor over 24 hours (Yang et al., 2015a) is not directly comparable to a 12.4% mass loss in T. castaneum over 60 days (Wang et al., 2020), because the experimental designs, measurement methods, larval densities, temperature conditions, and initial polymer molecular weights differ in ways that may dominate the apparent efficiency difference. Jiang et al. (2021a) compared PS degradation across T. molitor, G. melonella, and Z. atratus larvae under controlled identical conditions — one of the few studies to attempt such a controlled cross-species comparison — and their data illustrate that even under standardized conditions, the three species differ meaningfully in degradation rate and product profile.
What can be said with some confidence, despite these limitations, is that: (i) plastic degradation capacity is not confined to a single insect taxon but is distributed across Lepidoptera and Coleoptera at minimum; (ii) gut microbiome composition is substantially more diverse in plastic-fed larvae than in controls on conventional diets, consistent with the idea that plastic degradation products support a broader trophic base within the gut microbial community (Lou et al., 2020); (iii) the rate and extent of degradation are influenced by plastic type, molecular weight, and polymer branching structure in ways that reflect the differing enzymatic accessibility of different polymer chemistries (Yang et al., 2022); and (iv) dietary co-supplementation with conventional nutrients consistently improves degradation efficiency, larval survival, and microbiome diversity relative to plastic-only diets, a practical finding with direct relevance to any application design (Brandon et al., 2018; Yang et al., 2018, 2023).
It is also worth acknowledging a dimension of this literature that does not receive enough attention: the toxicological implications. Sanchez-Hernandez (2021) raised the concern that plastic degradation intermediates — oligomers, functionalized monomers, and adsorbed chemical additives released during depolymerization — may exert toxic effects on the insect hosts themselves, on the microbial communities within their guts, or on any organisms consuming plastic-fed insects (a relevant consideration if insect biomass is to be used as animal feed or fertiliser, as some proposals envision). The data here are thin, and the gap between degradation efficiency measurements and systematic toxicological characterization of end-products represents one of the most important outstanding needs in the field. Goveas et al. (2023) pointed to insect larvae gut microbiota as a potentially important tool for microplastic removal, but stressed that both the removal mechanisms and the fate of removal products require deeper investigation before practical applications can be responsibly designed.
3.7 Synthesis and Discussion: What This Body of Evidence Collectively Tells Us
Stepping back from the species-by-species and mechanism-by-mechanism detail, several themes emerge from this literature that deserve explicit synthesis.
The first, and perhaps most important, is that insect gut environments appear to function as naturally optimized bioreactor conditions for plastic polymer breakdown — not through any single overwhelming mechanism, but through the co-occurrence of multiple facilitating factors that, individually, are insufficient but collectively enable degradation at rates unachievable in open environmental conditions. Mechanical size reduction, substrate emulsification, host-derived salivary oxidases, and a dense, functionally diverse microbial community operating under physiologically controlled temperature, pH, and oxygen conditions — all present simultaneously in the insect gut lumen — constitute a degradation system of a kind that reductive single-mechanism analyses tend to miss. The practical implication is that efforts to engineer or exploit this biology for scale-up should probably focus on recreating these system conditions, rather than isolating individual enzymes or strains and expecting them to perform equivalently outside the gut context.
The second theme is one of genuine mechanistic uncertainty that the literature has not yet resolved. The relative contributions of host-intrinsic enzymatic activity (salivary oxidases, gut wall secretions) versus gut microbiome-derived enzymatic activity to overall degradation vary by species, by plastic type, and possibly by larval developmental stage in ways that have not been systematically mapped. The Kong et al. (2019) and Wang et al. (2022) findings suggesting partial microbiome independence in G. melonella are genuinely puzzling in this context, and the failure of antibiotic suppression experiments to cleanly separate host from microbial contributions in all systems reflects the deep physiological integration of these two components. More precise tools — germ-free larval rearing, targeted microbiome manipulation, and in vivo enzyme activity profiling — will be needed to disentangle these contributions.
Third, and related: the biosynthetic pathways governing production and regulation of plastic-degrading enzymes in gut microbial communities have not been characterized at a genomic or transcriptomic level in any of the species reviewed here. This is a significant absence. The discovery pipeline from gut microbiome to plastic-degrading enzyme to characterized biosynthetic pathway to rational engineering target — the pipeline that has worked well in antibiotic discovery and industrial enzyme biotechnology — has not been meaningfully entered for this system. Metagenomics of plastic-fed larval gut communities (see Bae et al., 2021; Peng et al., 2022 for early examples) begins to open this pipeline, but the analytical depth needed for biosynthetic pathway reconstruction has not yet been applied at scale.
The fourth and final theme is the scale-up question, which the literature largely sidesteps. Demonstrating that 100 larvae can consume 92 mg of PE in 12 hours is scientifically striking but industrially remote from the 370 million metric tons produced annually. Bridging that gap — whether through direct larval-based bioprocessing, enzyme extraction and immobilization, or microbial culture scale-up informed by gut microbiome discoveries — requires engineering, economic, and regulatory analysis that has barely begun. This review does not attempt to address those challenges directly, but it is worth naming them, because the gap between "fascinating biological discovery" and "practically applicable solution" is the gap that most of this literature has not yet seriously engaged.