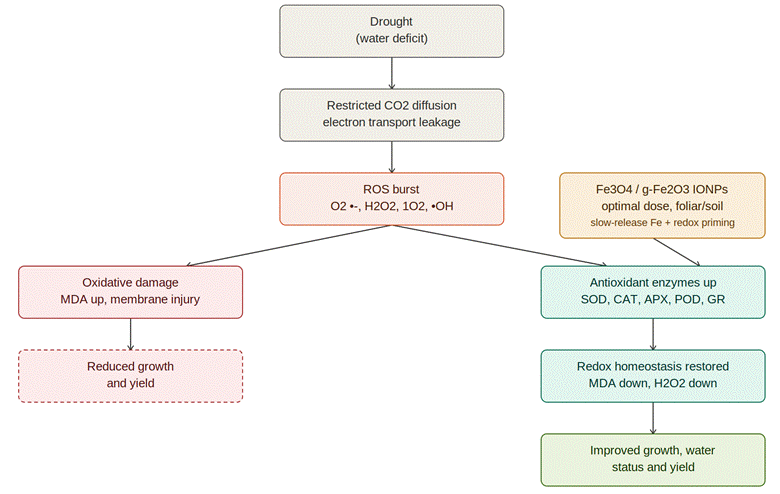
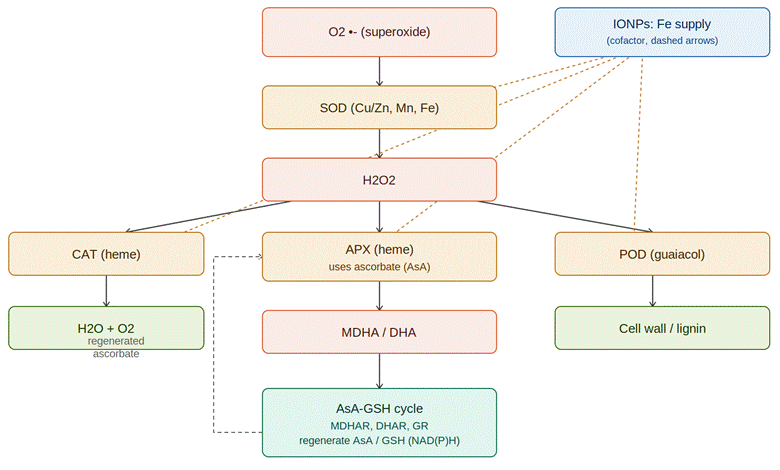
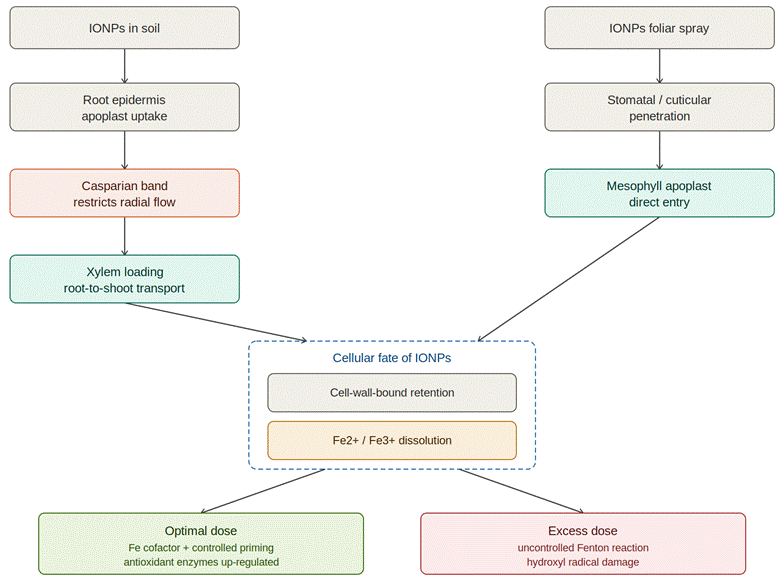
Bao, L., Liu, J., Mao, T., Zhao, L., Wang, D., & Zhai, Y. (2024). Nanobiotechnology-mediated regulation of reactive oxygen species homeostasis under heat and drought stress in plants. Frontiers in Plant Science, 15, Article 1418515. https://doi.org/10.3389/fpls.2024.1418515
Chandrashekar, H. K., Singh, G., Kaniyassery, A., Thorat, S. A., Nayak, R., Murali, T. S., & Muthusamy, A. (2023). Nanoparticle-mediated amelioration of drought stress in plants: A systematic review. 3 Biotech, 13, Article 336. https://doi.org/10.1007/s13205-023-03751-4
Choudhury, F. K., Rivero, R. M., Blumwald, E., & Mittler, R. (2017). Reactive oxygen species, abiotic stress and stress combination. The Plant Journal, 90(5), 856–867. https://doi.org/10.1111/tpj.13299
Das, K., & Roychoudhury, A. (2014). Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Frontiers in Environmental Science, 2, Article 53. https://doi.org/10.3389/fenvs.2014.00053
El-Zohri, M., Al-Wadaani, N. A., & Bafeel, S. O. (2021). Foliar sprayed green zinc oxide nanoparticles mitigate drought-induced oxidative stress in tomato. Plants, 10(11), Article 2400. https://doi.org/10.3390/plants10112400
Feng, Y., Kreslavski, V. D., Shmarev, A. N., Ivanov, A. A., Zharmukhamedov, S. K., Kosobryukhov, A., Yu, M., Allakhverdiev, S. I., & Shabala, S. (2022). Effects of iron oxide nanoparticles (Fe3O4) on growth, photosynthesis, antioxidant activity and distribution of mineral elements in wheat (Triticum aestivum) plants. Plants, 11(14), Article 1894. https://doi.org/10.3390/plants11141894
Foyer, C. H., & Shigeoka, S. (2011). Understanding oxidative stress and antioxidant functions to enhance photosynthesis. Plant Physiology, 155(1), 93–100. https://doi.org/10.1104/pp.110.166181
Gama, J. P. S., Campos, F. G., Riccardi, C. d. S., & Boaro, C. S. F. (2025). Iron oxide nanoparticles for photosynthetic recovery in iron-deficient 'Micro-Tom' tomato plants. Environments, 12(10), Article 346. https://doi.org/10.3390/environments12100346
Ghafariyan, M. H., Malakouti, M. J., Dadpour, M. R., Stroeve, P., & Mahmoudi, M. (2013). Effects of magnetite nanoparticles on soybean chlorophyll. Environmental Science & Technology, 47(18), 10645–10652. https://doi.org/10.1021/es402249b
Gill, S. S., & Tuteja, N. (2010). Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiology and Biochemistry, 48(12), 909–930. https://doi.org/10.1016/j.plaphy.2010.08.016
Harb, A., Awad, D., & Samarah, N. (2015). Gene expression and activity of antioxidant enzymes in barley (Hordeum vulgare L.) under controlled severe drought. Journal of Plant Interactions, 10(1), 109–116. https://doi.org/10.1080/17429145.2015.1033023
Hasanuzzaman, M., Bhuyan, M. H. M. B., Zulfiqar, F., Raza, A., Mohsin, S. M., Mahmud, J. A., Fujita, M., & Fotopoulos, V. (2020). Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants, 9(8), Article 681. https://doi.org/10.3390/antiox9080681
Javaid, A., Munir, N., Abideen, Z., Duarte, B., Siddiqui, Z. S., Haq, R., & Naz, S. (2025). The potential effects of nanoparticles in gene regulation and expression in mammalian, bacterial and plant cells — A comprehensive review. Plant Nano Biology, 11, Article 100145. https://doi.org/10.1016/j.plana.2025.100145
Khan, S., Akhtar, N., Rehman, S. U., & Jamil, M. (2023). Iron oxide nanoparticles: Plant response, interaction, phytotoxicity and defense mechanisms. In A. Husen (Ed.), Nanomaterials and nanocomposites exposures to plants (pp. 227–245). Springer. https://doi.org/10.1007/978-981-99-2419-6_11
Li, H., Wang, Z., Zhou, C., Wang, H., Chen, L., Yang, H., & Liu, D. (2025). Harnessing nanoparticles to enhance crop production under drought stress: A quantitative meta-analysis. Agricultural Water Management, 315, Article 109550. https://doi.org/10.1016/j.agwat.2025.109550
Li, X., Zhang, H., Tian, L., Huang, L., Liu, S., Li, D., & Song, F. (2015). Tomato SlRbohB, a member of the NADPH oxidase family, is required for disease resistance against Botrytis cinerea and tolerance to drought stress. Frontiers in Plant Science, 6, Article 463. https://doi.org/10.3389/fpls.2015.00463
Lu, K., Shen, D., Liu, X., Dong, S., Jing, X., Wu, W., Tong, Y., Gao, S., & Mao, L. (2020). Uptake of iron oxide nanoparticles inhibits the photosynthesis of the wheat after foliar exposure. Chemosphere, 259, Article 127445. https://doi.org/10.1016/j.chemosphere.2020.127445
Parvin, K., Hasanuzzaman, M., Mohsin, S. M., Nahar, K., & Fujita, M. (2024). Vanillic acid modulates antioxidant defense and methylglyoxal detoxification systems to combat drought stress in tomato seedlings. Plants, 13(22), Article 3114. https://doi.org/10.3390/plants13223114
Petrovic, I., Savic, S., Gricourt, J., Causse, M., Jovanovic, Z., & Stikic, R. (2021). Effect of long-term drought on tomato leaves: The impact on metabolic and antioxidative response. Physiology and Molecular Biology of Plants, 27(12), 2805–2817. https://doi.org/10.1007/s12298-021-01102-2
Quinet, M., Angosto, T., Yuste-Lisbona, F. J., Blanchard-Gros, R., Bigot, S., Martinez, J.-P., & Lutts, S. (2019). Tomato fruit development and metabolism. Frontiers in Plant Science, 10, Article 1554. https://doi.org/10.3389/fpls.2019.01554
Racuciu, M., Barbu-Tudoran, L., & Oancea, S. (2025). Evaluation of phytotoxicity and genotoxicity of TMA-stabilized iron-oxide nanoparticle in corn (Zea mays) young plants. Scientific Reports, 15, Article 18951. https://doi.org/10.1038/s41598-025-03872-1
Raza, A., Charagh, S., Abbas, S., Hassan, M. U., Saeed, F., Haider, S., Sharif, R., Anand, A., Corpas, F. J., Jin, W., & Varshney, R. K. (2023). Nano-enabled stress-smart agriculture: Can nanotechnology deliver drought and salinity-smart crops? Journal of Sustainable Agriculture and Environment, 2(3), 189–214. https://doi.org/10.1002/sae2.12061
Rehman, A., Khan, S., Sun, F., Peng, Z., Feng, K., Wang, N., Jia, Y., Pan, Z., He, S., Wang, L., Qayyum, A., Du, X., & Li, H. (2024). Exploring the nano-wonders: Unveiling the role of nanoparticles in enhancing salinity and drought tolerance in plants. Frontiers in Plant Science, 14, Article 1324176. https://doi.org/10.3389/fpls.2023.1324176
Rezayian, M., Niknam, V., & Arabloo, M. (2023). Iron nanoparticle regulate succinate dehydrogenase activity in canola plants under drought stress. Scientific Reports, 13, Article 9628. https://doi.org/10.1038/s41598-023-36105-4
Rizwan, H. M., Shafqat, U., Ishfaq, A., Batool, F., Mahmood, F., Su, Q., Yaseen, N., Raza, T., & Altihani, F. A. (2025). Identifying the phytotoxicity of biosynthesized metal oxide nanoparticles and their impact on antioxidative enzymatic activity in maize under drought stress. Plants, 14(7), Article 1075. https://doi.org/10.3390/plants14071075
Rizwan, M., Ali, S., Ali, B., Adrees, M., Arshad, M., Hussain, A., Zia ur Rehman, M., & Waris, A. A. (2019). Zinc and iron oxide nanoparticles improved the plant growth and reduced the oxidative stress and cadmium concentration in wheat. Chemosphere, 214, 269–277. https://doi.org/10.1016/j.chemosphere.2018.09.120
Sánchez-Rodríguez, E., Rubio-Wilhelmi, M. M., Cervilla, L. M., Blasco, B., Rios, J. J., Rosales, M. A., Romero, L., & Ruiz, J. M. (2010). Genotypic differences in some physiological parameters symptomatic for oxidative stress under moderate drought in tomato plants. Plant Science, 178(1), 30–40. https://doi.org/10.1016/j.plantsci.2009.10.001
Shahzad, R., Harlina, P. W., Khan, S. U., Koerniati, S., Hastilestari, B. R., Ningrum, R. A., Wahab, R., Djalovic, I., & Prasad, P. V. (2024). Iron oxide nanoparticles alleviate salt-alkaline stress and improve growth by modulating antioxidant defense system in cherry tomato. Journal of Plant Interactions, 19(1), Article 2375508. https://doi.org/10.1080/17429145.2024.2375508
Sharma, P., Jha, A. B., Dubey, R. S., & Pessarakli, M. (2012). Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. Journal of Botany, 2012, Article 217037. https://doi.org/10.1155/2012/217037
Soleymani, S., Piri, S., Aazami, M. A., & Salehi, B. (2025). Cerium oxide nanoparticles alleviate drought stress in apple seedlings by regulating ion homeostasis, antioxidant defense, gene expression, and phytohormone balance. Scientific Reports, 15, Article 11805. https://doi.org/10.1038/s41598-025-96250-w
Tao, Y., Gao, Q., Fan, X., Wu, H., & Shabala, S. (2026). From uptake to resilience: How metal-based nanoparticles can enhance plant drought tolerance. Plant and Soil, 519(2), 1121–1143. https://doi.org/10.1007/s11104-025-08230-8
Tombuloglu, H., Ercan, I., Alqahtani, N., Alotaibi, B., Bamhrez, M., Alshumrani, R., Turumtay, H., Ergin, I., Demirci, T., Ozcelik, S., Kayed, T. S., & Ercan, F. (2023). Impact of magnetic field on the translocation of iron oxide nanoparticles (Fe3O4) in barley seedlings (Hordeum vulgare L.). 3 Biotech, 13, Article 296. https://doi.org/10.1007/s13205-023-03727-4
Tombuloglu, H., Slimani, Y., Tombuloglu, G., Almessiere, M., & Baykal, A. (2019). Uptake and translocation of magnetite (Fe3O4) nanoparticles and its impact on photosynthetic genes in barley (Hordeum vulgare L.). Chemosphere, 226, 110–122. https://doi.org/10.1016/j.chemosphere.2019.03.075
Zhou, B., & Razzaq, S. (2025). Revolutionizing crop production with iron nanoparticles for controlled release of plant growth regulators and abiotic stress resistance. Plant Nano Biology. Advance online publication. https://doi.org/10.1016/j.plana.2025.100397
Zhou, R., Kong, L., Wu, Z., Rosenqvist, E., Wang, Y., Zhao, L., Zhao, T., & Ottosen, C.-O. (2019). Oxidative damage and antioxidant mechanism in tomatoes responding to drought and heat stress. Acta Physiologiae Plantarum, 41, Article 20. https://doi.org/10.1007/s11738-019-2805-1
Zhu, H., Han, J., Xiao, J. Q., & Jin, Y. (2008). Uptake, translocation, and accumulation of manufactured iron oxide nanoparticles by pumpkin plants. Journal of Environmental Monitoring, 10(6), 713–717. https://doi.org/10.1039/b805998e